CAPITULO 4
Anatomía funcional de las células
procarióticas y eucarióticas
A pesar de su complejidad y variedad todas las células vivas
pueden dividirse en dos grupos, las procariotas y las eucariotas, en base a su ultraestructura
tal como se aprecia al microscopio electrónico. Las plantas y los animales
están totalmente compuestos de células eucarióticas. En el mundo
microbiano las bacterias son procariotas. Los otros microbios celulares son eucariotas:
hongos (levaduras y mohos), protozoos y algas verdaderas.
Los virus, como elementos acelulares con ciertas propiedades de
tipo celular, no encajan en ningún esquema de organización de las células
vivas. Son partículas genéticas que se reproducen pero que son incapaces
de realizar las actividades químicas habituales de las células vivas.
El capítulo 12 trata de la estructura y actividad de los virus. Por el momento
nos concentraremos en el estudio de las células procarióticas y eucarióticas.
Los procariotas y los eucariotas son químicamente similares:
ambos contienen ácidos nucleicos, proteínas, lípidos y azúcares.
Utilizan los mismos tipos de reacciones químicas para metabolizar los nutrientes,
fabricar proteínas y almacenar energía. Es principalmente la estructura
de las paredes celulares, de las membranas y los orgánulos (estructuras
celulares especializadas que llevan a cabo funciones específicas) lo que distingue
a los procariotas de los eucariotas.
Las características clave para distinguir las células
procarióticas (lo que significa «prenúcleo» en griego)
son las siguientes:
1. Su material genético (DNA) no está encerrado dentro
de una membrana.
2. Carecen de otros orgánulos rodeados de membranas.
3. Su DNA no está asociado a proteínas de la clase
de las histonas (proteínas cromosómicas especiales que se hallan en
eucariotas).
4. Sus paredes celulares contienen casi siempre péptido
glucano, un polisacárido complejo.
5. Suelen dividirse por fisión binaria. En este proceso
se copia el DNA y la célula se divide en dos. La fisión binaria implica
menos estructuras y procesos que la mitosis y división celular de los eucariotas.
|
Objetivos
- Identificar las tres morfologías básicas de las
bacterias.
- Explicar las diferencias entre las paredes de las células
gram-positivas y gram-negativas.
- Describir las estructura, composición química y
funciones de la membrana citoplásmica.
- Definir difusión simple, ósmosis, difusión
facilitada, transporte activo y traslocación de grupos.
- Idenficar las funciones de las estructuras de las célula
procariótica.
- Comparar la estructura general de la célula, flagelos,
núcleo y ribosomas de procariotas y eucariotas.
- Explicar que es un orgánulo.
- Describir las funciones del retículo endoplásmico,
complejo de Golgi, mitocondrias, cloroplastos y lisosomas.
|

Las células eucarióticas (del griego «núcleo
verdadero») poseen estructuras lineales de DNA llamadas cromosomas que se encuentran
en el núcleo celular, el cual está separado del citoplasma por una
membrana nuclear. El DNA de los cromosomas eucarióticos está asociado
de forma constante a proteínas especiales cromosómicas, llamadas histonas
y no-histonas. Los eucariotas presentan también un aparato mitótico
(varias estructuras celulares que participan en un tipo de división nuclear
llamada mitosis) y diversos orgánulos, entre los que se encuentran las mitocondrias,
el retículo endoplásmico y a veces los cloroplastos. Revisaremos más
adelante las características y funciones particulares de estos orgánulos.
La celula procariática
Los miembros del mundo procariota comprenden un vasto y heterogéneo
grupo de organismos unicelulares. Este grupo incluye las eubacterias o bacterias
verdaderas y las arqueobacterias. Los millares de especies bacterianas se diferencian
por muchos factores, entre los que se cuentan la morfología, la composición
química (a menudo detectada por reacciones de tinción), los requerimientos
nutricionales y las actividades bioquímicas.
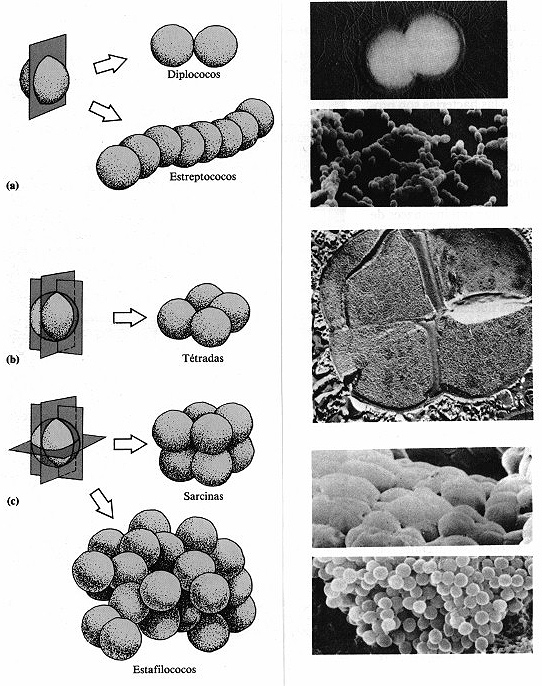
Figura 4-1. Disposiciones de los cocos. La disposición de las células
viene determinada por el número de planos por los que se divide cada célula.
A la izquierda aparecen los esquemas y a su derecha la correspondiente fotografía.
(a) La división según un plano da lugar a diplococos (15.000x)
y estreptococos (2.500x). (b) La división por dos planos origina tétradas
(25.000x). (c) Las sarcinas (16.000x) y los estafilococos (2.500x) son el
resultado de la división siguiendo tres planos.
Tamaño, morfología y disposición de las
células bacterianas
Se da una gran variedad de tamaños y morfologías
entre las bacterias. La mayoría de ellas oscilan entre 0,2 y 2,0 µm
de diámetro y presentan una de las tres morfologías básicas
siguientes: la esférica o de coco (que significa «baya»),
la de bastoncillo o bacilo y la espiral.
Los cocos suelen ser esféricos, pero pueden ser ovalados,
alargados o con un lado aplanado. Cuando los cocos se dividen para reproducirse pueden
permanecer unidos uno a otro. Los cocos que permanecen en parejas tras dividirse
se llaman diplococos (figura 4-1 a). Aquellos que se dividen en dos planos
y forman grupos de cuatro se conocen como tétradas (figura 4-1 b).
Los que se dividen por tres planos regulares y quedan divididos en grupos cúbicos
de ocho se llaman sarcinas (figura 4-1 c) y aquellos que se dividen siguiendo
planos al azar y forman racimos como los de uva o anchas láminas son estafilococos
(figura 4-1 c). Estas agrupaciones son frecuentemente útiles para la identificación
de algunos cocos.
Los bacilos se dividen solamente a través de su eje más
corto, de forma que hay menos agrupamientos de bacilos que de cocos. Los diplobacilos
aparecen en parejas tras la división (figura 4-2b) y los estreptobacilos
se presentan en cadenas (figura 4-2 y lámina A-7). Algunos bacilos tienen
el aspecto de cigarrillos, otros poseen extremos afilados como los cigarros puros.
Existen aún otros que son ovalados y se parecen tanto a los cocos que se llaman
cocobacilos (figura 4-2d). Sin embargo, la mayoría de los bacilos se
presentan de forma aislada (figura 4-2a).
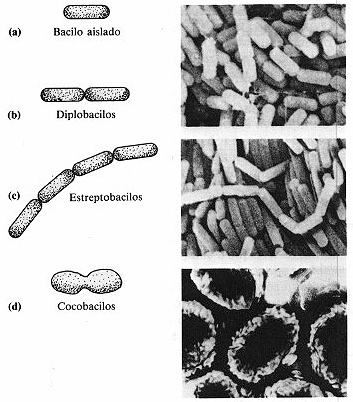
Figura 4-2. Bacilos. A la izquierda se muestran los esquemas y a la derecha sus
correspondientes fotografias. (a) Bacilos aislados. (b) Díplobacilos.
En la foto se aprecian algunas parejas de bacilos unidos que sirven de ejemplo de
diplobacilos (2.000x). (c) Estreptobacílos (2.000x). (d) Cocobacílos
(20.000x).
El término bacilo tiene dos significados en microbiología;
usado como hasta ahora se refiere a la morfología bacteriana, pero puede ser
también el origen de un nombre genérico. Por ejemplo, la bacteria Bacillus
anthracis es el agente etiológico del carbunco.
Las bacterias espirales pueden tener una o más vueltas;
nunca aparecen rectas. Los bacilos curvados en forma de coma se denominan vibrios
(figura 4-3 a). Otros, llamados espirilos, poseen una morfología helicoidal
característica, que recuerda un sacacorchos, con un cuerpo celular bastante
rígido (figura 4-3b). Hay aún otro grupo de bacterias espirales llamadas
espiroquetas (figura 4-3 c). A diferencia de los espirilos, que poseen flagelos,
las espiroquetas se mueven por medio de un filamento axial, parecido a un
flagelo pero que se encuentra dentro de una vaina externa flexible que rodea a la
bacteria. Como se verá más adelante, algunos géneros bacterianos
presentan morfologías y disposiciones más complejas.

Figura 4-3. Bacterias espirales. Esquema (izquierda) y sus correspondientes fotografías
(derecha) de (a) vibríos (4.800x), (b) espirilos (17.000x) y
(c) espiroquetas (4.000x).
Además de las tres morfologías básicas hay
también células en forma de estrella (género Stella)
y se han descubierto recientemente células planas, cuadrangulares (el
propuesto género Arcula) (figura 4-4).
La morfología de una bacteria viene determinada por la herencia.
Sin embargo, ciertas condiciones ambientales pueden alterar esa morfología
y cuando esto ocurre la identificación se hace aún más difícil.
Además algunas bacterias, como Rhizobium y Corynebacterium, son
genéticamente pleomórficas, lo que significa que pueden presentar muchas
morfologías.
En la figura 4-5 se muestra la estructura de una célula
procariótica (bacteriana) característica. Estudiaremos sus componentes
atendiendo al siguiente orden: (1) estructuras externas a la pared celular,
(2) pared celular y (3) estructuras internas a la pared celular.
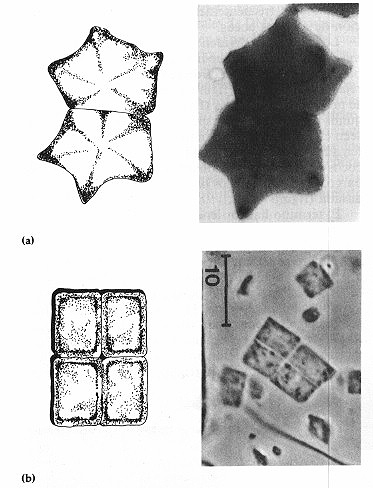
Figura 4-4. Células en forma de estrella y cuadrangulares. (a)
SteIIa (en forma de estrella) (36.000x). (b) Arcula (de forma
cuadrada) (1.700x).
Estructuras externas a la pared celular
Entre las estructuras externas a la pared celular procariótica
se encuentran el glucocálix, que puede tomar la forma de una cápsula
o de una capa mucilaginosa, los flagelos, los filamentos axiales y las fimbrias (pili).
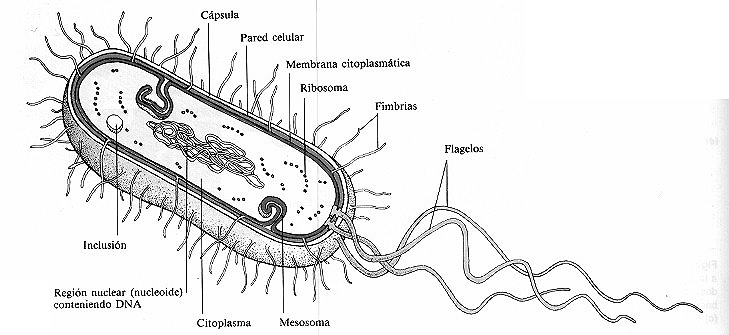
Figura 4-5. Estructura de una célula procariótica (bacteriana) típica.
En este esquema, un corte longitudinal revela las estructuras internas.
Glucocálix
Glucocálix es un término general empleado
para referirse a las sustancias que rodean a las células bacterianas (también
puede hablarse simplemente de pojímeros extracelulares). El glucocálix
es un polímero gelatinoso compuesto de polisacáridos, de polipéptidos,
o de ambos. Su complejidad química varía ampliamente entre las distintas
especies bacterianas. El material del glucocálix es viscoso (pegajoso) y en
su mayoría se forma en el interior de la células para ser excretado
a la superficie. Si esta sustancia está organizada y firmemente unida a la
pared celular el glucocálix se denomina cápsula (ver figura
14-2).
La presencia de cápsula puede determinarse mediante una
tinción negativa, como el método de la tinta china mencionado en el
capítulo 3 (ver figura 3-10a y lámina A-5). Si el material está
sin organizar y sólo débilmente adherido a la pared celular, el glucocálix
se describe como una capa mucilaginosa.
Las cápsulas son un importante mecanismo de virulencia (la
medida en la que un patógeno produce enfermedad) en algunas especies. Las
cápsulas protegen a menudo a las bacterias patógenas de la fagocitosis
por células del huésped. La fagocitosis, que se trata en el capítulo
15, es un proceso por el cual algunas células blancas sanguíneas
engloban y destruyen a los microbios (ver figura 15-8). Streptococcus pneumoniae
produce neumonía cuando sus células están protegidas por
una cápsula de polisacárido. Las cepas no capsuladas de S. pneumoniae
son fácilmente fagocitadas y no causan la enfermedad. Bacillus anthracis
produce una cápsula de ácido D-glutámico. Se recordará
del capitulo 2 que las formas D de los aminoácidos son poco corrientes. Como
B. anthracis es causa del carbunco sólo cuando presenta cápsula
parece que ésta le protege de la fagocitosis.
Otra función del glucocálix pegajoso es la adhesión
de la bacteria sobre distintas superficies con el fin de sobrevivir en su ambiente
natural. Mediante la adhesividad las bacterias pueden fijarse a superficies tan diversas
como las rocas de un torrente de aguas rápidas, las raíces de las plantas,
los dientes y tejidos humanos e incluso a otras bacterias. Streptococcus mutans;
una importante causa de caries dental, se adhiere a la superficie del diente
mediante un glucocálix. La cápsula de Klebsiella pneumoniae previene
la fagocitosis y permite a esta bacteria adherirse y colonizar el tracto respiratorio.
S. mutans puede utilizar su cápsula como fuente de
nutrientes cuando su reserva de energía es escasa, degradándola y empleando
sus azúcares. El glucocálix puede proteger a la célula de la
deshidratación. Además, su viscosidad puede inhibir el alojamiento
de nutrientes de la célula.
Flagelos
Un segundo componente estructural de algunas células procarióticas
son los flagelos (figura 4-5). Los flagelos (palabra que significa látigo)
son largos apéndices filamentosos que impulsan a la bacteria.
Los flagelos se presentan en las células bacterianas en
cuatro disposiciones diferentes (figura 4-6 a): monotricos (un solo flagelo),
anfitricos (un solo flagelo en cada extremo de la célula), lofotricos
(dos o más flagelos en uno o en ambos extremos de la célula) y
peritricos (flagelos distribuidos en toda la superficie celular).
El flagelo tiene tres partes fundamentales (figura 4-6 b). La región
más externa, el filamento, es constante en diámetro y contiene
la proteína globular (aproximadamente esférica) flagelina, dispuesta
en varias cadenas que forman una hélice alrededor de un núcleo hueco.
Las proteínas flagelares sirven para identificar algunas bacterias. El filamento
se une a un gancho ligeramente más grueso, formado por una proteína
distinta. La tercera porción del flagelo es el cuerpo basal, que le
sirve de anclaje a la pared celular y a la membrana citoplasmática.
El cuerpo basal se compone de un pequeño eje central insertado
en una serie de anillos. En las bacterias gram-negativas contiene dos pares de anillos.
Los externos se anclan en las distintas porciones de la pared celular y el par de
anillos internos lo hace en la membrana citoplasmática. En las bacterias gram-positivas
sólo está presente el par más interno. Como se verá en
este mismo capítulo, la estructura de los flagelos (y cilios) de las células
cucariotas es más compleja que la de las células cucariotas.
Las bacterias flageladas son móviles, es decir, tienen la
capacidad de moverse por sí mismas. Cada flagelo procariótico es un
rotor helicoidal, semirrígido, que empuja a la célula al girar a partir
del cuerpo basal. La rotación del flagelo puede ser en el sentido de las agujas
de un reloj o en el contrario, a lo largo de su eje. En cambio, los flagelos eucarióticos
tienen un movimiento ondulante. El movimiento de un flagelo procariótico es
la consecuencia de la rotación de su cuerpo basal y es similar al giro del
eje de un motor eléctrico. Al rotar el flagelo forma un remolino que desplaza
el agua y propulsa a la bacteria (figura 4-6c). Aunque las bases exactas químicas
y mecánicas de este «motor» biológico no están totalmente
aclaradas, se sabe que depende de la continua generación de energía
por parte de la célula.
Las células bacterianas pueden alterar la velocidad y dirección
de rotación de sus flagelos y son, por tanto, capaces de exhibir distintas
formas de movilidad (figura 4-6c). Cuando una bacteria se mueve en una dirección
durante un período de tiempo se llama a este mo vimiento una «carrera»,
o se dice que «nada». Las «carreras» se ven interrumpidas
por cambios periódicos de dirección, bruscos y al azar, llamados «tumbos»
o «volteretas». Después prosigue su carrera. Los tumbos están
provocados por la inversión del sentido de rotación de sus flagelos.
Algunas especies de bacterias dotadas de un gran número de flagelos (por ejemplo,
Proteus) pueden «invadir» un medio de cultivo, exhibiendo un crecimiento
rápido en forma de ondas sobre él (ver lámina A-14).

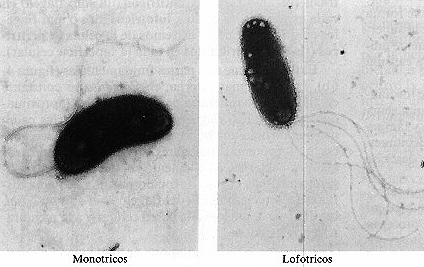
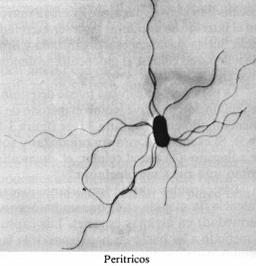
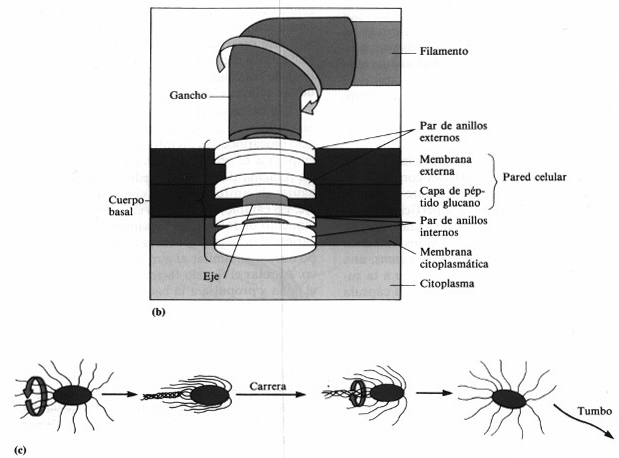
Figura 4-6. Flagelos. (a) Los cuatro
tipos básicos de disposición flagelar: anfitricos (SpiriIIum ostreae
3.500 x ) monotrícos (víbrio marino 21.000x), lofotricos (Ectothiorhodospira
moblis, 2.000x) y peritrícos (Proteus vulgaris, 4.200x). (b)
Elementos y fijación del flagelo de una bacteria gram-negativa. (c)
Formas de movilidad bacteriana mostrando una «carrera» y un «tumbo».
Una ventaja de la movilidad es que permite a una bacteria moverse
hacia un ambiente favorable o alejarse de uno adverso. La aproximación o alejamiento
de una bacteria con respecto a un estímulo determinado se llama taxis.
Tales estímulos pueden ser químicos (quimiotaxis) o luminosos
(fototaxis). Las bacterias móviles poseen receptores en diversas localizaciones,
como en la pared celular o debajo de ella. Estos receptores recogen estímulos
químicos, como oxígeno, ribosa o galactosa. En respuesta al estímulo
la información es transmitida a los flagelos. Si la señal quimiotáctica
es positiva es un atractante, la bacteria se mueve hacia el estímulo
con muchas carreras y pocos tumbos. Si la señal quimiotáctica es negativa,
un repelente, aumenta la frecuencia de los tumbos y la bacteria se aleja del
estímulo (véase el recuadro de la pág. 80).
Filamentos axiales
Las espiroquetas son un grupo de bacterias que poseen una estructura
y movilidad exclusivas. Una de las espiroquetas mejor conocidas es Treponema pallidum,
el agente etiológico de la sífilis. Las espiroquetas se mueven
por medio de filamentos axiales que emergen de ambos extremos de la célula
y la rodean en espiral por debajo de la vaina (figura 4-7). Los filamentos axiales
poseen una estructura similar a la de los flagelos y están anclados en un
extremo de la espiroqueta. La rotación de los filamentos hace que la célula
helicoidal y rígida gire en dirección opuesta avanzando como un sacacorchos.
Las células eucarióticas no poseen filamentos axiales.
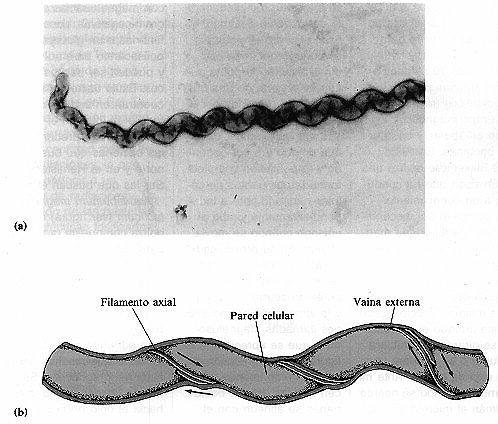
Figura 4.7. Filamentos axiales. (a) Microfotografía de la espiroqueta
Leptospira mostrando un filamento axial (14.000x). (b) Esquema de los
filamentos axiales que rodean parcialmente una espiroqueta.
Fimbrias
Las fimbrias, o pili (en singular pilus), son
apéndices en forma de vellosidades unidos a las células bacterianas
de forma muy parecida a los flagelos, pero son considerablemente más cortas
y delgadas (ver figura 4-5). Como los flagelos, las fimbrias están compuestas
por una proteína (llamada pulina) dispuesta helicoidalmente
alrededor de un núcleo central. Las fimbrias pueden presentarse en los extremos
de la célula bacteriana o estar uniformemente distribuidas sobre toda la superficie.
Pueden darse desde unas pocas a varios cientos de fimbrias por célula (figura
4-8). Muchas bacterias gramnegativas poseen fimbrias. Las células cucarióticas
no las poseen.
Hay fimbrias de dos tipos, con funciones diferentes. El primer
tipo, las fimbrias comunes, permiten a la célula adherirse a las superficies,
incluyendo las de otras células. Esta función es similar a la del glucocálix.
Las fimbrias presentes en la bacteria Neisseria gonorrhoeae, el agente causal
de la gonorrea, ayudan al microorganismo a colonizar las mucosas. Una vez que ha
tenido lugar la colonización, la bacteria es capaz de producir la enfermedad.
Cuando las fimbrias están ausentes (a causa de una mutación genética)
no puede tener lugar la colonización y no se produce la enfermedad.
Un segundo tipo de fimbria, llamadas pelos (o pili)
sexuales, facilitan la unión de las células bacterianas
previa a la transferencia de DNA desde una célula a la otra. Estos pili
se estudian con detalle en el capítulo 8.
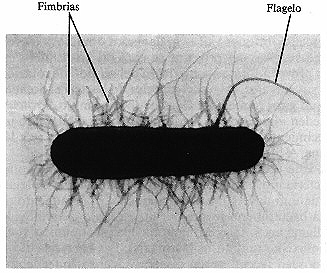
Figura 4-8. Fimbrias. Esta célula de E. coli aparece erizada de
fimbrias comunes (22.000x). Obsérvese el flagelo.
[Página
Siguiente]