|
Imanes microbianos
Durante cerca de 100 años los biólogos han sospechado
que los pájaros son capaces de utilizar el campo magnético terrestre
para orientarse durante el vuelo. En los años 70 se demostró en algunos
pájaros la magnetotaxis (respuesta a un campo magnético), así
como en abejas y en algunas bacterias. En 1975 R. P. Blakemore realizó una
observación que ha conducido a un conocimiento más completo del mecanismo
responsable de la magnetotaxis. Describió la presencia de bacterias muy móviles
en barros ricos en sulfuros que «...se movían en la misma dirección
geográfica aunque el microscopio se girara, se cambiara de lugar o se cubriera
con una caja de cartón. Ante mi asombro, cuando se acercó un imán
al microscopio, los cientos de células móviles ¡giraron instantáneamente
y se alejaron precipitadamente del extremo del imán! Eran siempre atraídas
por el extremo del imán que atraía también la punta que señala
el norte de una brújula y eran repelidas por el extremo opuesto. Su velocidad
era muy rápida, del orden de 100 µm por segundo y la población
completa formada por cientos de células nadando libre e independientemente
viraba al unísono cuando se movía el imán en su proximidad»*.
Las bacterias magneto-tácticas sintetizan magnetita, óxido
de hierro (Fe3O4) y lo almacenan en inclusiones llamadas magnetosomas,
que se aprecian en la figura. Los magnetosomas actúan como imanes y hacen
que las células bacterianas se alineen con el campo geomagnético terrestre.
Todas las bacterias con magnetosomas son gram-negativas, poseen fimbrias o un glucocálix
de polisacárido para adherirse y pueden ser bacilos o cocos. Estas bacterias
se encuentran en sedimentos en todo el munto. En el Hemisterio Norte predominan las
bacterias que buscan el norte y en el Hemisferio Sur las que buscan el sur Aquaspirillum
magnetotacticum (ver figura) metaboliza compuesta orgánicos para obtener
carbono y energía y requiere un ambiente microaerófilo. Se ha observado
que cuando A. magnetotacticum es separado del lugar donde está adherido
nada a lo largo de lo que resultan ser líneas geomagnéticas. Se mueve
descendiendo, bien hacia el polo norte o bien hacia el polo sur hasta alcanzar un
lugar de fijación adecuado.
Se está actualmente estudiando cómo sintetizan estas
bacterias la magnetita y aún no está clara la función de este
compuesto en el metabolismo de la célula. La formación de ion férrico
(Fe3+) a partir de ion ferroso (Fe2+) podría proporcionar ATP a la célula, pero se ha estimado
que la cantidad de ATP es tan pequeña que la magnetita debe tener otra función.
In vitro los magnetosomas pueden descomponer peróxido de hidrógeno,
que se forma en la célula en presencia de oxigeno y suele ser degradado por
la enzima catalasa antes de que alcance concentraciones tóxicas. Los investigadores
especulan sobre la posibilidad de que los magnetosomas protejan in vivo a
la célula de la acumulación de peróxido de hidrógeno.
La síntesis y función de los magnetosomas interesa
a los microbiólogos para completar nuestro conocimiento sobre las células
y puede proporcionar un modelo para explicar la formación de óxidos
de hierro similares en pájaros, insectos y otros animales. La ecología
de los microbios magnéticos puede suministrar además una herramienta
a los científicos que estudian el campo magnético terrestre. Hay evidencias
que sugieren que el campo magnético terrestre puede haberse invertido o alterado
varias veces. El examen de la orientación de los fósiles de bacterias
magnetotáctícas preservadas en rocas o sedimentos puede arrojar luz
sobre el movimiento relativo de los continentes y los polos magnéticos. Las
bacterias magnetotácticas también podrían ayudarnos a comprender
la evolución. Normalmente el campo magnético de la Tierra repele una
parte de los gases ionizados que emanan continuamente del Sol. Durante una inversión
magnética la Tierra es golpeada por una cantidad mayor de estas radiaciones,
lo que podría conducir a un aumento de radiaciones. La orientación
de las bacterias magnetotácticas fosilizadas en relación a los mutantes
fósiles podría ayudarnos a saber si las inversiones magnéticas
afectan a la evolución.
*Reproducido con permiso del Annual Review of Microbiology volumen
36. © 1982 por Annual Reviews Inc. y Richard R Blakemore. (Richard R
Blakemore, 1982. «Magnetotactic Bacteria». Annual Review of Microbiology
36: 217-238).
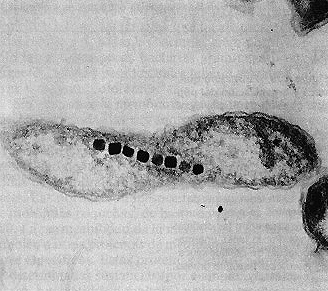
Aquaspirillum magnetotacticum mostrando una cadena de magnetosomas. Es
también visible la membrana externa de la pared gram-negativa (30.000x).
|
La pared celular
La pared celular de las células bacterianas es una estructura
compleja, semirrigida, responsable de la morfología característica
de la célula. La pared celular recubre la frágil membrana citoplasmática
y la protege a ella y a las partes internas de la célula de los cambios adversos
del medio ambiente (ver figura 4-5). Casi todos los procariotas tienen pared celular.
La función primordial de la pared celular es prevenir la
ruptura de la célula bacteriana cuando su presión osmótica interna
es mayor que la del medio externo. Sirve también de punto de anclaje a los
flagelos, ayuda a mantener la morfología de la célula y provoca síntomas
de enfermedad en algunas especies. A medida que aumenta el volumen de la célula
la membrana citoplasmática y la pared celular recrecen en concordancia. Clinicamente
la pared celular es importante por ser el lugar de acción de algunos antibióticos.
Aunque algunos eucariotas, como plantas, algas y hongos, poseen
paredes celulares, éstas difieren químicamente de las de los procariotas,
su estructura es más simple y son menos rígidas.
Composición y características
La pared celular bacteriana está formada por una red macromolecular
llamada péptido glucano (o mureina). El péptido glucano
es un mucopolisacárido formado por unidades repetidas de un disacárido
unido a cadenas de cuatro o cinco aminoácidos. Los monosacándos que
lo forman, N-acetil-glucosamina (NAG) y ácido N-acetil-murámico (NAM)
(de murus, pared), derivan de la glucosa y llevan unidos aminoácidos.
Las fórmulas estructurales del NAG y NAM aparecen en la figura 4-9.
Los distintos componentes del péptido glucano se ensamblan
en la pared celular de la siguiente manera. La N-acetil-glucosamina y el ácido
N-acetil-murámico aparecen alternados formando cadenas, constituyendo cada
una de ellas un «esqueleto» azucarado (figura 4-10). Hay de 10 a 65 moléculas
de azúcar en cada cadena. A cada molécula de ácido N-acetil-murámico
se une una cadena lateral de tetrapéptido, formada por cuatro aminoácidos.
Estos aminoácidos se presentan en forma D y L alternadas (ver figura 2-16),
algo exclusivo del péptido glucano puesto que los aminoácidos que se
hallan en las proteínas son siempre de la forma L.
Las cadenas laterales adyacentes de tetrapéptidos pueden
unirse una a otra directamente o a través de un puente peptídico
cruzado, que consta de uno a cinco aminoácidos.
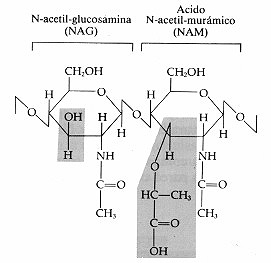
Figura 4-9. N-acetil-glucosamina (NAG) y ácido N-acetil-murámico (NAM)
unidos como aparecen en el péptido glucano. La unión que establecen
se denomina de tipo ß-1,4. Las zonas coloreadas señalan las diferencias
entre ambas moléculas.
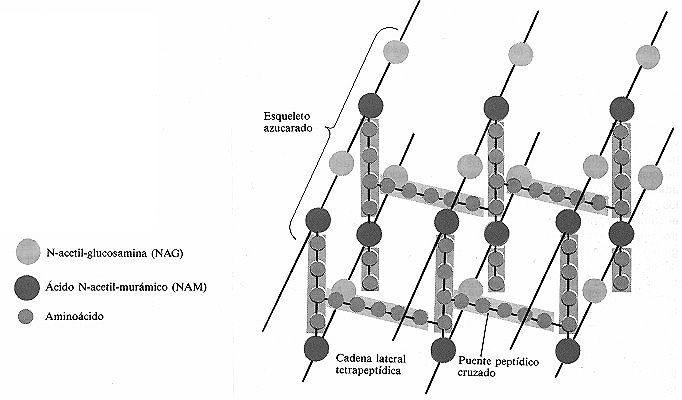
Figura 4-10. Estructura química y organización del péptido glucano
de Staphylococcus aureus. La frecuencia de los puentes peptídicos
cruzados y el número de aminoácidos que contienen varían según
la especie bacteriana. Además de los puentes existentes entre las cadenas
de cada lámina de péptido glucano hay otros entre las distintas láminas.
Por lo tanto, una macromolécula de péptido glucano puede ser bastante
gruesa.
En la mayoría de las bacterias gram-positivas la pared celular
consta de varias capas de péptido glucano conectadas por cadenas peptidicas
laterales y puentes cruzados, tal como aparece en la figura 4-1 0. Esta disposición
da lugar a una estructura muy rígida. Las capas de péptido glucano
son considerablemente más espesas en las bacterias gram-positivas que en las
gramnegativas (figura 4-11 a). Las paredes celulares de muchas bacterias gram-positivas
contienen ácidos teicoicos, que hacen posible la identificación
de estas bacterias por métodos inmunológicos, como se describirá
en el capítulo 16. Los ácidos teicoicos están formados principalmente
por un alcohol (glicerol o ribitol) y fosfato y están unidos a las capas de
péptido glucano o a la membrana citoplamática. Debido a su carga negativa
(de los grupos fosfato) los ácidos teicoicos pueden controlar la entrada y
salida de cationes (iones positivos) de la célula uniéndose a ellos.
También pueden tener una función en el crecimiento celular. Cuando
la pared celular crece se precisa de unas enzimas llamadas autolisinas. Estas
enzimas separan componentes de la pared para permitir la inserción de nuevas
subunidades. Los ácidos teicoicos regulan la actividad de las autolisinas,
evitando una rotura extensiva de la pared y la posible lisis. Finalmente, los ácidos
teicoicos pueden estar implicados en el almacenamiento de fósforo. Las paredes
celulares de las bacterias ácido-alcohol resistentes, como Mycobacterium,
están formadas por péptido glucano y hasta un 60 % de lípidos.
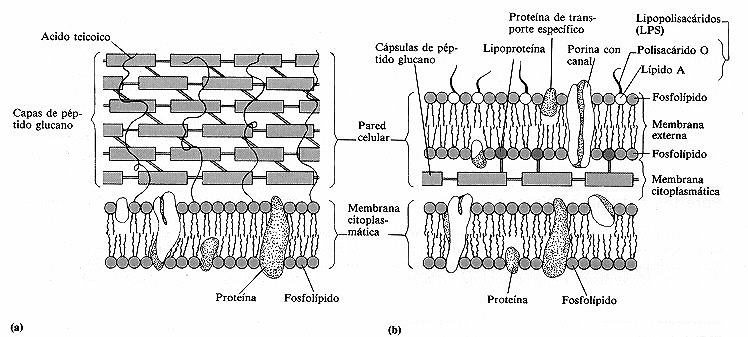
Figura 4-11. Comparación entre las estructuras y los componentes de las paredes
celulares de bacterias gram-positivas (a) y gram-negativas (b).
Las bacterias gram-negativas también contienen péptido
glucano, pero en una proporción muy pequeña y no poseen en absoluto
ácidos teicoicos. El péptido glucano se encuentra en el espacio
periplásmico, espacio existente entre la membrana citoplasmática
y la membrana externa y está unido covalentemente a las lipoproteínas
de esta última membrana. El espacio periplásmico tiene la consistencia
de un gel y contiene una elevada concentración de enzimas degradativas y proteínas
de transporte. Al contener tan sólo una pequeña cantidad de péptido
glucano las paredes celulares de las bacterias gram-negativas son más sensibles
a la ruptura mecánica.
La capa de péptido glucano de las bacterias gramnegativas
está rodeada de una membrana externa compuesta de lipoproteinas, lipopolisacáridos
(LPS) y fosfolípidos (figura 4-11 b). Mientras que las lipoproteinas de la
membrana citoplasmática están constituidas por proteínas unidas
covalentemente a lípidos, las de la membrana externa de la pared celular constan
de proteínas unidas no covalentemente a lípidos. La membrana externa
tiene varias funciones especializadas. Su fuerte carga negativa es un factor importante
para evadir la acción de los fagocitos y del complemento, dos componentes
de las defensas del huésped (que se tratan con detalle en el capítulo
15).
La membrana externa supone además una barrera para sustancias
como los antibióticos (por ejemplo, la penicilina), lisozima (enzima que puede
romper las paredes celulares de todas las bacterias gram-positivas pero sólo
las de unas pocas bacterias gram-negativas), detergentes, metales pesados, sales
biliares, enzimas digestivas y algunos colorantes.
Sin embargo, la membrana externa no supone una barrera para todas
las sustancias presentes en el medio ambiente puesto que los nutrientes deben atravesarla
para mantener el metabolismo celular. Parte de la permeabilidad de la membrana externa
se debe a las proteínas de membrana llamadas porinas que forman canales.
Las porinas son inespecíficas y permiten el paso de moléculas pequeñas,
de hasta un peso molecular de 800. La permeabilidad de la membrana externa se debe
también a otras proteínas de membrana llamadas receptores específicos.
Estas proteínas permiten sólo el paso de determinadas sustancias (por
ejemplo, vitamina B12, hierro, nucleótidos y maltosa) y sirven también
de lugar de fijación de virus y bacteriocinas (proteínas producidas
por algunas bacterias que inhiben o matan a especies muy relacionadas).
El componente lipopolisacarídico (LPS) de la membrana externa
es responsable de dos características importantes de las bacterias gram-negativas.
La parte polisacarídica del LPS se compone de azúcares, llamados polisacáridos
O, que son antígenos y ayudan a distinguir especies de bacterias gram-negativas
(por ejemplo,
Salmonella) por métodos serológicos. Este
papel es comparable al de los ácidos teicoicos en las células grampositivas.
La porción lipídica del LPS, llamada lipido A, se conoce como
endotoxina por ser tóxico cuando se libera en el torrente sanguíneo
del huésped. Produce fiebre y choque. La naturaleza e importancia de ésta
y de otras toxinas se discutirá en el capítulo 14.
La tabla 4-1 compara algunas características de las bacterias
gram-positivas y gram-negativas.
TABLA 4-1. Comparación de algunas
características de bacterias gram-positivas y gram-negativas.
|
Carácteristicas
|
Gram-positivas
|
Gran-negativas
|
| Tinción de Gram |
Retienen el cristal violeta y se tiñen de morado |
Se decoloran y toman el colorante de contraste (safranina), tiñéndose
de rosa. |
| Capa de péptido glucano |
Gruesa (varias capas) |
Delgada (una capa). |
| Contenido en lipopolisacáridos (LPS) |
Virtualmente ninguna |
Alta |
| Contenido en lípidos y lipoproteína |
Bajo (las bacterias ácido-alcohol resistentes tienen lípidos
unidos al péptido glucano) |
Alto (debido a la presencia de membrana externa). |
| Acidos teicoicos |
Presentes en muchas de ellas |
Ausentes. |
| Espacio periplásmico |
Ausente |
Presente. |
| Membrana externa |
Ausente |
Presente. |
| Estructura de los flagelos |
2 anillos en el cuerpo basal |
4 anillos en el cuerpo basal. |
| Producción de toxinas |
Principalmente exotoxinas |
Principalmente endotoxinas. |
| Resistencia a la rotura física |
Alta |
Baja. |
| Sensibilidad de la pared celular a la lisozima |
Altá |
Baja (requiere un tratamiento previo para desestabilizar la membrana
externa). |
| Marcada |
Mucho menos marcada. |
| Sensibilidad a penicilina y sulfamidas |
| Sensibilidad a estreptomicina, cloranfenicol y tetraciclinas |
Mucho menos marcada |
Marcada. |
| Inhibición por colorantes básicos |
Marcada |
Mucho menos marcada. |
| Sensibilidad a detergentes aniónicos |
Marcada |
Mucho menos marcada. |
| Resistencia a la azida sódica |
Marcada |
Mucho menos marcada |
| Resistencia a la desecación |
Alta |
Baja. |
Paredes celulares atípicas
Entre los procariotas se encuentran de forma natural células
que no poseen paredes o que presentan muy poco material de pared. Entre ellas se
incluyen los miembros del género Mycop1asma y microorganismos relacionados.
Los micoplasmas son las bacterias más pequeñas que se conocen que puedan
crecer y reproducirse fuera de las células vivas del huésped. Como
no poseen paredes celulares atraviesan la mayoría de los filtros bacterianos
y fueron confundidas al principio con virus. Sus membranas citoplasmáticas
son únicas entre las bacterias por contener lípidos llamados esteroles,
que se cree que contribuyen a protegerla de la lisis por presión osmótica.
Las arqucobacterias tienen paredes celulares compuestas por azúcares y proteínas
y nunca presentan péptido glucano.
Otras células bacterianas atípicas son las formas
L (nombradas así en honor al Instituto Lister, donde fueron descubiertas).
Se trata de pequeñas bacterias muantes defectivas en su pared celular. Algunos
compuestos químicos y antibióticos como la penicilina pueden inducir
a muchas bacterias a producir formas L. Aunque algunas formas L pueden revertir a
su forma bacteriana original, otras son estables. Las formas L tienden a contener
justo el material de la pared celular suficiente para prevenir su lisis cuando se
encuentran en soluciones diluidas.
Recuérdese del capítulo 3 que el fundamento de la
tinción de Gram está relacionado con la estructura y composición
de la pared celular, por lo que sería útil repasarlo ahora.
Lesiones en la pared celular
Las sustancias químicas que lesionan las paredes celulares
bacterianas, o que interfieren con su síntesis, no suelen ser dañinas
para las células del huésped animal porque su pared tiene una composición
química diferente de la de las células eucarióticas.
Por consiguiente, la síntesis de la pared celular es el
blanco de acción de algunos fármacos antimicrobianos. Una forma por
la que puede ser lesionada la pared celular es por la exposición a la enzima
llamada lisozima. Esta enzima aparece de forma natural en algunas células
eucariotas y es un componente de las lágrimas, mucosidad y saliva. Es particularmente
activa sobre los principales componentes de la pared celular de la mayoría
de bacterias gram-positivas, haciéndolas vulnerables a la ruptura o lisis.
La lisozima cataliza la hidrólisis de los enlaces que unen las moléculas
de azúcar en las cadenas polisacarídicas del péptido glucano.
Esta acción es análoga a cortar con un soplete los soportes de acero
de un puente. La pared celular de las bacterias gram-positivas es destruida casi
totalmente por la lisozima. Sin embargo, el contenido celular que permanece rodeado
por la membrana citoplasmática puede conservarse intacto si no tiene lugar
la lisis osmótica (ver más adelante); esta célula sin pared
se denomina protoplasto. Los protoplastos son típicamente esféricos
y capaces aún de realizar su metabolismo.
Cuando se aplica la lisozima de forma similar a las células
gram-negativas su pared no suele destruirse en la misma medida que las de las gram-positivas,
conservándose además parte de la membrana externa. En este caso el
contenido celular, la membrana citoplasmática y el resto de la capa externa
de la pared, constituyen un esferoplasto, que es también una estructura
esférica. Para que la lisozima actúe sobre las células gramnegativas
es necesario tratarlas antes con ácido etiléndiamino tetracético
(EUTA), compuesto que debilita las células iónicas de la membrana externa
y la altera, permitiendo el acceso de la lisozima al péptido glucano.
Los protoplastos y esferoplastos explotan en agua destilada o en
soluciones muy diluidas de sal o azúcar porque las moléculas de agua
del fluido que las rodea difunden rápidamente al interior, agrandando la célula
que tiene un porcentaje de agua mucho más bajo. Esta rotura se conoce como
lisis osmótica y se discutirá con detalle un poco más
adelante.
Algunos antibióticos, como la penicilina, destruyen las
bacterias al interferir con la formación de los puentes peptídicos
del péptido glucano, impidiendo, por tanto, la formación de la pared
celular funcional. La mayoría de las bacterias gram-negativas no son tan sensibIes
a la penicilina porque la membrana externa de las bacterias gram-negativas forma
una barrera que impide la entrada de la penicilina y otras sustancias. Los microorganismos
sin pared celular, como los micoplasmas y las formas L, son resistentes a los antibióticos
que inhiben la síntesis de la pared celular. Las bacterias gramnegativas son
bastante sensibles a los antibióticos ß-lactámicos distintos
de la penicilina. En el capítulo 19 se trata con más detalle de los
antibióticos.
Estructuras internas a la pared celular
Hasta aquí hemos estudiado la pared celular procariótica
y las estructuras externas a ella. Miraremos ahora el interior de la célula
procariótica para examinar la estructura y función de la membrana citoplasmática
y de otros componentes del citoplasma de la célula.
Membrana citoplasmática
La membrana ciloplasmática es una delgada estructura
que se extiende por dentro de la pared celular encerrando el citoplasma de la célula
(ver figura 4-5). La membrana citoplasmática procariótica está
formada principalmente por fosfolípidos (figura 2-13), que son el componente
mayoritario y por proteínas. Las membranas citoplasmáticas eucarióticas
contienen además azúcares y esteroles, como el colesterol. Por su carencia
de esteroles las membranas citoplasmáticas de procariotas son menos rígidas
que las eucariotas. Una excepción la constituyen las procariotas sin pared
llamados micoplasmas, cuya membrana contiene esteroles.
Estructura
En las fotografías al microscopio electrónico tanto
las membranas celulares procarióticas como eucarióticas aparecen como
bicapas: se observan dos líneas oscuras separadas por un espacio claro (figura
4-12 a). Las moléculas de fosfolípidos están dispuestas en dos
filas paralelas formando una bicapa fosfolipídica (figuras 4-12b y
c). Cada molécula de fosfolípido consta de una cabeza polar formada
por un grupo fosfato y glicerol que es hidrófila (afín al agua) y soluble
en agua y una cola apolar compuesta por ácidos grasos que son hidrófobos
(repelen el agua) e insolubles en agua (figura 4-12 a). Los extremos polares se sitúan
en las dos superficies de la bicapa fosfolipídica y los extremos apolares
en el interior de la bicapa.
Las moléculas de proteína de la membrana pueden disponerse
de diferentes maneras (figura 4-12c). Algunas, llamadas proteinas superficiales,
son extraídas fácilmente de la membrana por tratamiento químico
y se cree que yacen sobre la superficie interna o externa de la membrana. Pueden
actuar como enzimas que catalizan reacciones químicas, como «andamios»
de soporte y como mediadores en los cambios morfológicos de la membrana durante
el movimiento. Otras, llamadas proteínas integrales, pueden extraerse
de la membrana sólo después de desorganizar la bícapa. Se cree
que penetran total o parcialmente en la membrana. Algunas de estas proteínas
contienen canales que permiten la entrada y salida de sustancias en la célula.
Estudios realizados por físicos y biólogos han demostrado que las moléculas
de fosfolípidos y proteínas no están estáticas en la
membrana sino que parecen moverse con bastante libertad dentro de ellas. Este movimiento
se asocia muy probablemente a muchas de las funciones realizadas por la membrana
citoplasmática. La disposición dinámica de los fosfolípidos
y proteínas se conoce como el modelo del mosaico fluido (véase
el recuadro del capítulo 2, pág. 42).
Funciones
La función más importante de la membrana citoplasmática
es actuar como una barrera selectiva a través de la cual entran y salen sustancias
de la célula. En esta tarea las membranas citoplasmáticas son selectivamente
permeables (también se las denomina a veces semipermeables). Este
término indica que determinadas moléculas e iones atraviesan la membrana
mientras que otros no pueden. La permeabilidad de la membrana depende de varios factores.
Las moléculas grandes (como las proteínas) no pueden atravesarla porque
son mayores que los canales de las proteínas integrales. Las moléculas
más pequeñas (como agua, aminoácidos y algunos azúcares
sencillos) suelen pasar fácilmente a través de la membrana.
Los iones penetran tan sólo muy lentamente. Las sustancias
que se disuelven fácilmente en lípidos (como oxígeno, dióxido
de carbono y moléculas orgánicas apolares) entran y salen con mucha
más facilidad que otras puesto que la membrana se compone principalmente de
fosfolípidos. El movimiento de materiales a través de la membrana citoplasmática
depende también de las moléculas transportadoras que se describirán
en breve.
Las membranas citoplasmáticas son también importantes
en la degradación de nutrientes y en la producción de energía.
Las membranas citoplasmáticas de las bacterias contienen enzimas capaces de
catalizar las reacciones químicas que rompen las sustancias nutritivas y préducen
ATP. En algunas bacterias se encuentran pigmentos y enzimas implicados en la fotosíntesis
(la conversión de la energía luminosa en energía química)
localizados en invaginaciones de la membrana citoplasmática que se extienden
por el citoplasma. Estas capas de membranas se llaman cromatóforos o tilacoides
(figura 4-13).
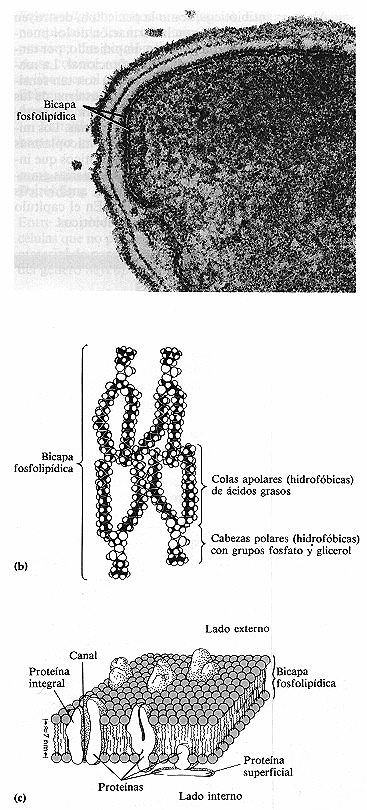
Figura 4-12. Membrana citoplasmática. Tal como se dispone en la bicapa
fosfolipídica. (a) Fotografía al microscopio electrónico
de la bicapa fosfolipidíca que forma la membrana citoplasmática de
la bacteria Bacillus brevis (175.000x). Pueden verse las capas de la pared
celular por fuera de la membrana. (b) Modelos compactos de varias moléculas.
(c) Dibujo de una membrana mostrando la bicapa de fosfolípidos y proteínas.
Mesosomas
Las membranas citoplasmáticas de las bacterias contienen
a menudo uno o más plegamientos grandes, irregulares, denominados mesosomas
(ver figura 4-5). Generalmente los mesosomas se ven asociados a la zona nuclear
o próximos al lugar de división de la célula. Los mesosomas
no están presentes en las células eucariotas.
Aunque la función exacta de los mesosomas es desconocida
podrían jugar un papel en la reproducción y el metabolismo. Cuando
se divide una célula bacteriana (fisión binaria) se forma una pared
llamada septo transverso y el material genético de la célula
parental se reparte entre las dos células hijas idénticas (ver figura
4-14). Los mesosomas podrían iniciar la formación del septo transverso
y unir el DNA bacteriano a la membrana citoplasmática. Los mesosomas podrían
ayudar también a separar el DNA en cada célula hija después
de la fisión binaria. Algunos científicos creen que los mesosomas pueden
tener un función en la generación de ATP y en la secreción de
enzimas al exterior de la célula. También la habilidad de las células
para concentrar nutrientes podría verse incrementada por la presencia de mesosomas,
puesto que sus plegamientos aumentan la superñcie de la membrana citoplasmática.
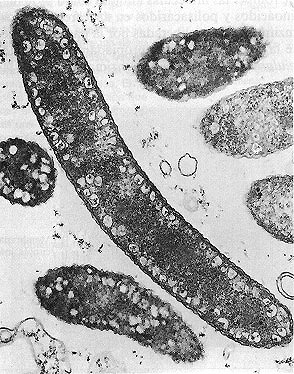
Figura 4-13. Cromatóforos. En esta totografia al microscopio electrónico
de transmisión de Rhodospirillum rubrum, una bacteria roja del azutre
pueden verse estructuras membranosas donde tiene lugar la fotosíntesis (30.000x).
Hay un debate considerable sobre la estructura y función
de los mesosomas. Se ha sugerido incluso que los mesosomas podrían ser artefactos
(estructuras que aparecen en las preparaciones microscópicas a causa del método
de preparación). En otras palabras, los mesosomas podrían no ser estructuras
reales de las células vivas.
Destrucción de la membrana citoplasmática por
agentes antimicrobianos
Como la membrana citoplasmática es vital para la célula
bacteriana no es sorprendente que sea el lugar de acción de varios agentes
antimicrobianos. Además de los compuestos que dañan la pared celular
y dejan por tanto expuesta la membrana a lesiones indirectas, muchas sustancias alteran
específicamente las membranas plasmáticas. Entre estos compuestos se
incluyen ciertos alcoholes y derivados de amonio cuaternario que se utilizan como
desinfectantes. Al desorganizar los fosfolípidos de membrana, un grupo de
antibióticos conocidos como polimixinas provocan la pérdida
de componentes intracelulares y consiguientemente la muerte celular. Este mecanismo
se discute en el capitulo 19.
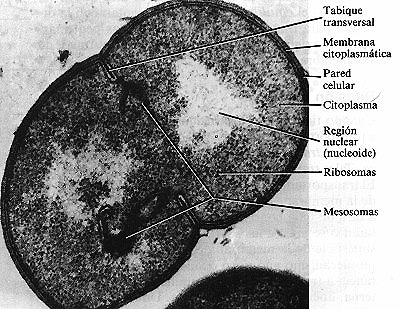
Figura 4-14. Tabique transversal en una célula en división de Sporosarcina
aureae. Pueden verse dos mesosomas que se extienden por el citoplasma a partir
de los bordes internos del tabique (72.000x).
Movimiento de sustancias a través de membranas
Cuando la concentración de una sustancia es mayor en un
lado de una membrana que en el otro, existe un gradiente (diferencia) de
concentraciones. Si la sustancia puede atravesar la membrana se moverá
hacia el lado donde esté más diluida, hasta que se igualen las concentraciones
o hasta que otras fuerzas detengan su movimiento.
Tanto en las células procarióticas como eucarióticas
las sustancias atraviesan las membranas citoplasmáticas por dos clases de
procesos: pasivos y activos. En los procesos pasivos las sustancias cruzan la membrana
desde una zona de alta concentración a otra de baja concentración (siguiendo
el gradiente de concentración), sin ningún gasto de energía
(ATP) por parte de la célula. Ejemplos de procesos pasivos son la difusión
simple, la ósmosis y la difusión facilitada. En los procesos activos
la célula debe consumir energía (ATP) para mover sustancias desde zonas
de baja concentración a otras de concentración más elevada.
Ejemplos de procesos activos son el transporte activo y la traslocación de
grupos.
Difusión simple
La difusión simple es el movimiento neto de moléculas
o iones de una zona de alta concentración a otra de baja concentración
(figura 4-15). Este movimiento se mantiene hasta que las moléculas o iones
estén distribuidos homogéneamente; en este momento se dice que se ha
alcanzado el equilibrio. Las células cuentan con la difusión
para transportar algunas moléculas pequeñas, como el oxígeno
y el dióxido de carbono, a través de sus membranas celulares.
Difusión facilitada
Cuando tiene lugar la difusión facilitada la sustancia
a transportar (por ejemplo, glucosa) se combina con una proteína transportadora
en la membrana citoplasmática. Tales transportadores se llaman a veces
permeasas. El transportador puede conducir a la sustancia a través
de la membrana desde una zona de alta concentración a otra de baja concentración.
Las proteínas transportadoras se unen a las sustancias que transportan en
la superficie de la membrana citoplasmática y mediante un mecanismo aún
poco conocido conducen a la sustancia a través de la membrana hasta la superficie
interna, liberádola en el citoplasma. Un mecanismo propuesto para la difusión
facilitada consiste en que la proteína transportadora permanece en su lugar,
pero experimenta un cambio conformacional que transporta a la sustancia de un lado
a otro de la membrana (figura 4-16). La difusión facilitada es similar a la
difusión simple ya que la sustancia se mueve siguiendo el gradiente de concentración
y la célula no gasta energía. El proceso se diferencia por el uso de
transportadores.
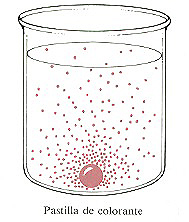
Figura 4-15. El fundamento de la difusión simple. Las moléculas
de colorante de la pastilla difunden en el agua desde una zona de alta concentración
de colorante hacia zonas de baja concentración.
En algunos casos las moléculas que necesita la bacteria
son demasiado grandes para ser transportadas al interior de la célula por
los métodos descritos. No obstante, la mayoría de las bacterias producen
enzimas capaces de romper las moléculas grandes como proteínas en aminoácidos
y polisacáridos en azúcares sencillos. Tales enzimas, que son liberadas
por la bacteria al medio que la rodea, se llaman apropiadamente enzimas extracelulares.
Una vez que las enzimas degradan las moléculas grandes las subunidades
producidas son transportadas por permeasas al interior de la célula. Por ejemplo,
transportadores específicos recuperan las bases del DNA, como la purina, guanina,
del medio extracelular y las llevan al citoplasma de la célula.
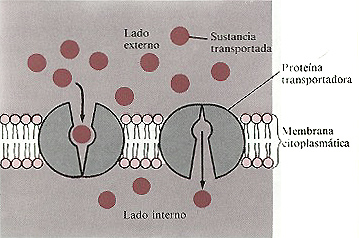
Figura 4-16. Difusión facilitada. Las proteinas transportadoras de la
membrana conducen moléculas a través de ésta de una zona de
alta concentración a otra de baja (siguiendo el gradiente de concentración).
La molécula transportadora sufre probablemente un cambio en su configuración
para facilitar el paso de la sustancia. Este proceso no requiere ATP
Osmosis
La ósmosis es el movimiento neto de moléculas de
solvente a través de una membrana selectivamente permeable, desde una zona
en la que las moléculas están fuertemente concentradas a otra de concentración
inferior. En los sistemas vivos el solvente fundamental es el agua.
La ósmosis puede ser demostrada con el aparato que se esquematiza
en la figura 4-17. Una bolsa de celofán, que es una membrana selectivamente
permeable, se llena con una solución acuosa coloreada de sacarosa al 20%.
La abertura de la bolsa de celofán se cierra con un tapón de goma atravesado
por un tubo de vidrio. La bolsa de celofán se coloca entonces en un vaso que
contiene agua destilada. Inicialmente las concentraciones de agua a cada lado de
la membrana son distintas: a causa de las moléculas de sacarosa la concentración
de agua es menor dentro de la bolsa de celofán que fuera. Para eliminar es
~ diferencia el agua se mueve desde el vaso (donde su concentración es mayor)
hacia el interior de la bolsa de celofán (donde su concentración es
menor).
No hay movimiento de azúcar hacia fuera de la bolsa de celofán
porque éste es impermeable a las moléculas de azúcar; las moléculas
de sacarosa son demasiado grandes para atravesar los poros de la membrana. A medida
que el agua penetra en la bolsa de celofán la solución de azúcar
se va diluyendo y como la bolsa está expandida al máximo por el incremento
del volumen de agua, ésta comienza a subir por el tubo de vidrio. Con el tiempo
el agua que se ha acumulado en la bolsa y en el tubo ejerce una presión hacia
abajo que fuerza a las moléculas de agua a salir de la bolsa de celofán
y volver al vaso. Este movimiento de agua a través de una membrana semipermeable
produce una presión llamada presión osmótica. Presión
osmótica es la presión necesaria para impedir el movimiento de
agua pura (sin solutos) hacia una solución que contiene algún soluto.
En otras palabras, la presión osmótica es la presión necesaria
para detener el flujo de agua a través de una membrana selectivamente permeable
(celofán). Cuando las moléculas de agua entran y salen de la bolsa
con la misma velocidad se ha alcanzado el equilibrio.
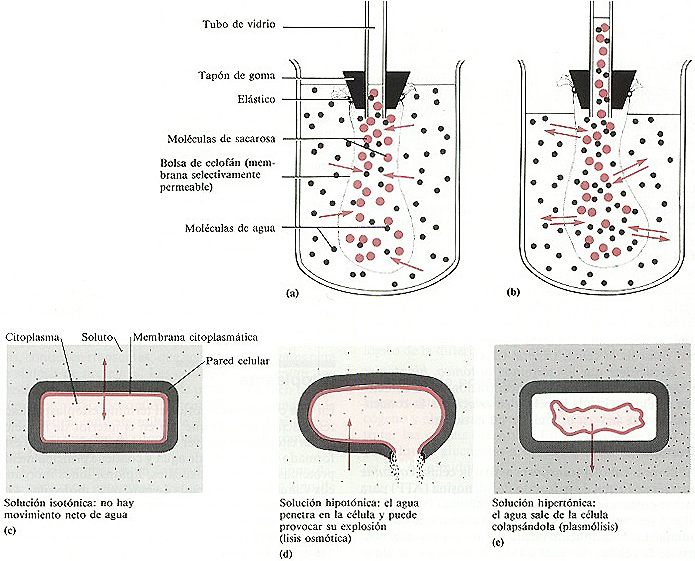
Figura 4-17. El fundamento de la ósmosis. (a) Situación
al comenzar el experimento. Las moléculas de agua comienzan a moverse del
vaso al interior de la bolsa siguiendo el gradiente de concentración. (b)
Situación de equilibrio. La presión osmótica ejercida por la
solución de la bolsa empuja las moléculas de agua de su interior hacia
el vaso, equilibrando la velocidad con que entra agua en la bolsa. El peso final
de la solución que hay en el tubo de vidrio en (b) es una medida de
la presión osmótica. (c)-(e) Efecto de diversas soluciones sobre
las células bacterianas.
Una célula bacteriana puede ser sometida a tres tipos de
soluciones osmóticas: isotónicas, hipotónicas o hipertónicas.
Una solución isotónica (Iso significa igual) es una en
la cual la concentración total de solutos es la misma a ambos lados de la
membrana. El agua entra y sale de la célula con la misma velocidad (no hay
movimiento neto) el contenido celular está en equilibrio con la solución
que hay en el exterior de la pared celular (figura 4-17c).
Se ha mencionado antes que la lisozima y algunos antibióticos
dañan la pared celular bacteriana causando la rotura o lisis de la célula.
Esta rotura tiene lugar porque el citoplasma bacteriano suele contener una concentración
tan alta de solutos que cuando la pared es debilitada o eliminada penetra agua en
la célula por ósmosis. La pared celular lesionada (o eliminada) no
puede contener la hinchazón de la membrana citoplasmática y ésta
revienta. Esto es un ejemplo de lisis osmótica causada por inmersión
en una solución hipotónica. Una solución hipotónica
(hipo significa menor o por debajo) en el exterior de la célula es
un medio cuya concentración de soluto es menor que la que hay en el interior
de la célula. La mayoría de las bacterias viven en soluciones hipotónicas
y su hinchazón es contenida por la pared celular. Las células con una
pared celular débil, como las bacterias gram-negativas, pueden sufrir lisis
osmótica por una entrada excesiva de agua (figura 4-17d).
Una solución hipertónica (hiper significa
por encima) es un medio con una concentración de solutos superior a la que
tiene la célula. La mayoría de las células bacterianas al ser
colocadas en una solución hipertónica se arrugan y colapsan porque
el agua abandona por ósmosis la célula (figura 4-17c).
Recuérdese que los términos isotónico,
hipotónico e hipertónico describen la concentración
de las soluciones en el exterior de la célula en relación a
la concentración que hay en el interior de la misma.
Transporte activo
Al llevar a cabo el transporte activo la célula consume
energía en forma de trifosfato de adenosina (ATP) para mover sustancias
a través de la membrana citoplasmática. Este movimiento suele ser del
exterior al interior, aunque la concentración pueda ser mucho más alta
dentro de la célula. Al igual que la difusión facilitada el transporte
activo depende de la presencia de proteínas transportadoras en la membrana
citoplasmática (figura 4-16). Parece haber distintos transportadores para
cada sustancia transportada o para cada grupo de sustancias estrechamente relacionadas.
Traslocación de grupos
En el transporte activo la sustancia que atraviesa la membrana
no es alterada por el proceso de transporte. En la traslocación de grupos,
una forma especial de transporte activo que tiene lugar exclusivamente en procariotas,
la sustancia es alterada químicamente durante su transporte a través
de la membrana. Una vez que la sustancia ha sido alterada y se halla dentro de la
célula la membrana citoplasmática se hace impermeable a ella, de forma
que permanecen en el interior de la célula. Esto es importante porque una
célula puede así acumular diversas sustancias aunque se encuentren
en bajas concentraciones fuera de la célula. La traslocación de grupos
requiere energia, sumimstrada por el fosfoenolpiruvato, un compuesto de fosfato de
alta energía.
Un ejemplo de traslocación de grupo es el transporte del
azúcar glucosa, que se utiliza a menudo en medios de cultivos para bacterias.
Durante su transporte por una proteína específica a través de
la membrana se añade un grupo fosfato a la molécula de glucosa. La
energía necesaria para la adición de fosfato es proporcionada por el
fosfoenolpiruvato. La forma fosforilada de la glucosa no puede ser transportada al
exterior y es utilizable en las rutas metabólicas de las células.
La difusión simple y la difusión facilitada son mecanismos
útiles para el transporte de sustancias al interior de las células
cuando su concentración es mayor en el medio externo. Sin embargo, cuando
una célula bacteriana se encuentra en un ambiente en el cual los nutrientes
están en bajas concentraciones la célula debe utilizar el transporte
activo y la translocación de grupos para acumular las sustancias necesarias.
Las células eucarióticas pueden utilizar dos procesos
adicionales de transporte activo llamados fagocitosis y picnocitosis. Ambos se describen
en apartados posteriores.
[Página
Anterior] [Página
Siguiente]