
Citoplasma
En una célula procariótica el término citoplasma hace referencia a todo lo que hay en el interior de la membrana citoplasmática (ver figura 4-5). El citoplasma está formado por un 80% de agua y contiene principalmente proteínas (enzimas), azúcares, lípidos, iones inorgánicos y muchos compuestos de bajo peso molecular. Los iones inorgánicos están presentes en el citoplasma en concentraciones mucho más altas que en la mayoría de los medios. El citoplasma es espeso, semitransparente y elástico. Las principales estructuras que se encuentran en el citoplasma son DNA, partículas llamadas ribosomas y depósitos de reserva llamados inclusiones.
El citoplasma procariótico carece de algunas características del citoplasma eucariótico, como un citoesqueleto y corrientes citoplasmáticas. Estas características se describirán más adelante.
Región nuclear
La reglón nuclear, o nucicoide, de una célula bacteriana (ver figura 4-5) contiene una única molécula circular, larga, de DNA bicatenario: el cromosoma bacteriano (ver figura 8-2a). Esta es la información genética de la célula, su DNA, portador de toda la información necesaria sobre la estructura y funciones celulares.
A diferencia de los cromosomas de las células eucarióticas, los cromosomas bacterianos no están rodeados por una cubierta nuclear. Los nucleoides pueden ser esféricos, alargados o en forma de halterio.
Las bacterias contienen a menudo, además del cromosoma bacteriano, pequeñas moléculas circulares de DNA bicatenario llamadas plásmidos (véase el factor F en la figura 8-23a). Se trata de elementos genéticos extracromosómicos, es decir, que no tienen conexión con el cromosoma bacteriano principal y que se replican de forma autónoma. Los plásmidos suelen contener de 5 a 100 genes, que no suelen ser cruciales para el crecimiento de la bacteria bajo las condiciones ambientales normales y pueden ser adquiridos o perdidos sin riesgo para la célula. Bajo determinadas condiciones, sin embargo, los plásmidos son ventajosos para la célula. Los plásmidos pueden albergar genes que determinen actividades como la resistencia a antibióticos, tolerancia a metales tóxicos, producción de toxinas y síntesis de enzimas. Los plásmidos pueden ser transferidos de una bacteria a otra.
Ribosomas
Todas las células, tanto eucarióticas como procarióticas, contienen ribosomas, que actúan como lugares para la síntesis de proteínas. Las células que presentan una elevada velocidad de síntesis de proteínas tienen un número mayor de ribosomas. El citoplasma de una célula procariótica contiene miles de estas pequeñísimas estructuras, que dan al citoplasma un aspecto granuloso (ver figura 4-5).
Los ribosomas están formados por dos subunidades, cada una compuesta por proteínas y un tipo de RNA llamado RNA ribosómico (RNAr). Los ribosomas procarióticos difieren de los eucarióticos en el número de proteínas y moléculas de RNA que contienen y son algo más pequeñas y menos densas que los ribosomas de las células eucarióticas. A causa de ello los ribosomas procarióticos se denominan ribosomas 70S y los de las células eucarióticas se conocen como ribosomas 80S. La letra S corresponde a unidades Svedberg, que indican la velocidad relativa de sedimentación durante la centrifugación a velocidad ultraelevada. La velocidad de sedimentación depende del tamaño, peso y morfología de la partícula. Las subunidades de un ribosoma 70S son una subunidad pequeña 30S que contiene una molécula de RNAr y una subunidad grande 50S que contiene dos moléculas de RNAr.
Varios antibióticos, como la estreptomicina, la neomicina y las tetraciclinas, ejercen sus efectos antimicrobianos inhibiendo la síntesis de proteínas en los ribosomas. Gracias a las diferencias entre los ribosomas procarióticos y eucarióticos pueden ser destruidas las bacterias por el antibiótico sin afectar a la célula eucariótica hospedadora.
Inclusiones
Dentro del citoplasma de las células procarióticas (y eucarióticas) se encuentran varios tipos de depósitos de reserva conocidos como inclusiones. Algunas inclusiones son comunes a una amplia variedad de bacterias mientras que otras están limitadas a un pequeño numero de especies y sirven, por lo tanto, como un criterio para la identificación. Entre las inclusiones bacterianas más notables se encuentran las siguientes:
Corpúsculos metacromáticos
Estas inclusiones se tiñen de rojo con algunos colorantes azules, como el azul de metileno y se conocen también como gránulos de volutina. Se trata de una forma de reserva de fosfato inorgánico (polifosfato) que puede utilizarse en la síntesis de ATP. La volutina se forma generalmente en células que crecen en ambientes ricos en fosfatos. Los corpúsculos metacromáticos se encuentran en algas, hongos y protozoos, así como en bacterias. Estos gránulos son bastante grandes y característicos en Corynebacierium diphtheriae, el agente etiológico de la difteria, por lo que tienen valor diagnóstico.
Gránulos de polisacáridos
Estas inclusiones están formadas típicamente por glucógeno y almidón y puede demostrarse su presencia al tratar las células con yodo. En presencia de yodo los gránulos de glucógeno aparecen marrón rojizo y azules los de almidón.
Inclusiones lipídicas
Las inclusiones lipídicas aparecen en varias especies de Mycobacierium, Bacillus, Azotobacter, Spirillum y otros géneros. Un lípido hallado corrientemente como material de reserva y exclusivo de las bacterias es el polímero de ácido poli-ß-hidroxi-butírico. Las inclusiones lipídicas se ponen de manifiesto utilizando colorantes liposolubles, como los colorantes de Sudán.
Gránulos de azufre
Ciertas bacterias, conocidas como ´bacterias del azufreª, que pertenecen al género Thiobacillus, obtienen energía al oxidar azufre y compuestos de azufre. Estas bacterias pueden depositar gránulos de azufre en el interior de la célula, donde le sirven como reserva de energía.
Carboxisomas
Son inclusiones poliédricas o exagonales que contienen la enzima ribulosa-l, 5-difosfato carboxilasa. Las bacterias que utilizan dióxido de carbono como única fuente de carbono necesitan esta enzima para la fotosíntesis. Entre las bacterias que contienen carboxisomas se encuentran las bacterias nitrificantes, las cianobacterias y las bacterias del azufre.
Vacuolas de gas
Son cavidades huecas que se encuentran en muchos procariotas acuáticos, incluyendo cianobacterias, bacterias fotosintéticas anoxigénicas y halobacterias. Cada vacuola está formada por filas de varias vesículas gaseosas individuales, que son cilindros huecos cubiertos por proteínas. La función de las vacuolas de gas es mantener la flotabilidad, de modo que las células puedan permanecer en el agua a la profundidad adecuada para recibir suficiente cantidad de oxígeno, luz y nutrientes.
Véase en el recuadro de la página 80 una discusión sobre las inclusiones de las bacterias magnetotácticas.
Endosporas
Cuando se han agotado los nutrientes esenciales, o cuando no disponen de agua, algunas bacterias gram-positivas, como las pertenecientes a los géneros Clostridium y Bacillus, forman células especializadas ´latentesª llamadas endosporas. Las endosporas, excusivas de las bacterias, son cuerpos deshidratados con una gruesa pared o capas adicionales. Se forman en el interior de la membrana celular y son liberadas al medio ambiente, donde pueden sobrevivir a temperaturas extremadas, falta de agua y exposición a muchos compuestos tóxicos y radiaciones.
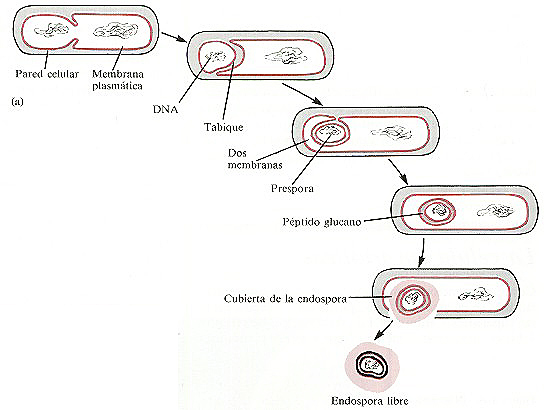
El proceso de la formación de endosporas dentro de la célula vegetativa (parental) se conoce como esporulación o esporogénesis (figura 4-18a y b). No está claro qué sucesos bioquímicos desencadenan este proceso. En la primera fase observable de la esporogénesis el crecimiento de una invaginación de la membrana citoplasmática llamada septo esporal aísla una pequeña porción del citoplasma y un cromosoma bacteriano recién replicado. El septo esporal se transforma en una membrana doble que rodea al cromosoma y al citoplasma. Esta estructura totalmente encerrada dentro de la célula original se conoce como prespora. Entre las dos membranas se depositan capas espesas de péptido glucano y a continuación se forma una gruesa cubierta de la espora de proteína alrededor de la membrana más externa. Es esta cubierta la responsable de la resistencia de las endosporas a muchos compuestos químicos agresivos.
El diámetro de la endospora puede ser el mismo, más pequeño o mayor que el de la célula vegetativa. Dependiendo de la especie la localización de la endospora puede ser terminal (en un extremo), subterminal (cerca de un extremo) o central, dentro de la célula vegetativa (figura 4-18c). Cuando la endospora madura la pared de la célula vegetativa se disuelve (se lisa) y la endospora queda libre.
La mayoría del agua presente en el citoplasma de la prespora se ha eliminado al completarse la esporogénesis y las endosporas no llevan a cabo reacciones metabólicas. El centro altamente deshidratado de la endospora contiene solamente DNA, pequeñas cantidades de RNA, ribosomas, enzimas y algunas moléculas pequeñas importantes. Entre estas últimas se incluye una cantidad sorprendentemente grande de un ácido orgánico llamado ácido dipicolinico, que va acompañado por una gran cantidad de ion calcio. Estos componentes celulares serán esenciales para reiniciar más tarde el metabolismo.
Las endosporas pueden permanecer latentes durante largos períodos de tiempo, incluso cientos de años y volver al estado vegetativo por un proceso denominado germinación. La germinación es desencadenada por lesiones físicas o químicas de la cubierta de la endospora. Entonces las enzimas de la endospora rompen las capas que la rodean, penetra agua y se reemprende el metabolismo. Como una célula vegetativa forma una sola endospora, que tras germinar se convierte en una célula, en las bacterias la esporogénesis no es una forma de reproducción; no hay un aumento en el número de células.
Las endosporas son importantes desde el punto de vista clínico y de la industria alimentaria porque son resistentes a procesos que matan normalmente a las células vegetativas. Tales procesos incluyen calentamiento, congelación, desecación, uso de compuestos químicos y de radiaciones. Mientras que la mayoría de las células vegetativas son destruidas por temperaturas que superen los 70 °C, las endosporas pueden sobrevivir una hora o más en agua hirviendo. Las bacterias formadoras de esporas constituyen un problema en la industria alimentaria porque tienen posibilidades de sobrevivir en caso de cualquier defecto en el tratamiento y porque cuando pueden crecer algunas especies producen toxinas y enfermedades. Los métodos especiales utilizados para el control de microorganismos productores de endosporas se discuten en el capítulo 7.
Como se señaló en el capítulo 3, las endosporas son difíciles de teñir y debe utilizarse un colorante especial conjuntamente con calor. Corrientemente se utiliza la tinción de Schaeffer-Fulton para endosporas.
Una vez examinada la anatomía funcional de las células procarióticas haremos lo mismo con las células cucarióticas.
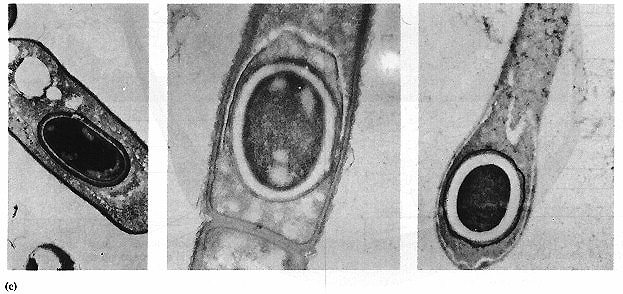
Figura 4-18. Endosporas. (a) Esporogénesis, el proceso de formación de endosporas. (b) Fotografía al microscopio electrónico de una endospora de Bacillus sphaericus. (c) Distintas localizaciones de las endosporas. Izquierda: Endospora central de Bacillus megateríum (40.500x); Centro: endospora subterminal de B. antrhacis (38.000x); derecha: endospora terminal, redonda, en el extremo de una célula de Clostridium tetani (14.000x aproximadamente).
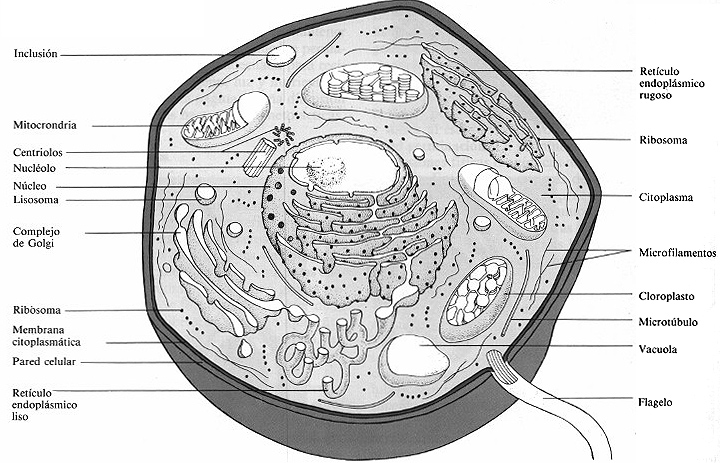
Figura 4-19. Diagrama muy esquemático de una célula eucariótica con todos los orgánulos posibles.
La célula eucariótica
Como se menciono antes, entre los organismos eucarióticos se encuentran algas, protozoos, hongos, plantas superiores y animales. La célula eucariótica (figura 4-19) es característicamente mayor y de estructura más compleja que la célula procariótica. Comparando la estructura de una célula procariótica que aparece en la figura 4-5 con la de la célula eucariótica podemos apreciar las diferencias que hay entre los dos tipos de células.
Brevemente las células eucarióticas contienen orgánulos rodeados de membranas; las células procarióticas no. Los orgánulos son estructuras especializadas que llevan a cabo funciones específicas. Aunque ambos tipos de células realizan las mismas funciones básicas las células procarióticas no las tienen localizadas en orgánulos específicos. El material genético (DNA) de las células procarióticas no está rodeado de una membrana ni asociado de forma regular a proteínas. El material genético de las células eucarióticas está rodeado de una membrana, organizado en cromosomas y estrechamente asociado a histonas y otras proteínas.
El estudio de la anatomía de la célula eucariótica seguirá de forma paralela el de la correspondiente a la célula procariótica, comenzando con las estructuras que se extienden mas alla del cuerpo de la célula. Al final de la discusión se resumirá en la tabla 4-2 las diferencias principales entre las células procarióticas y eucarióticas.
Flagelos y cilios
Muchos tipos de células eucarióticas poseen proyecciones que le sirven para la locomoción celular o para mover sustancias a lo largo de la superficie de la célula. Estas proyecciones contienen citoplasma y están rodeadas por la membrana citoplasmática. Si las proyecciones son escasas y largas en relación al tamaño de la célula se denominan flagelos. Cuando son numerosas y cortas, con aspecto de vellos, se conocen como cilios. Tanto los flagelos como los cilios constan de nueve pares de microtúbulos que rodean a un par interno central. Las algas euglenoides utilizan un flagelo para la locomoción mientras que los protozoos, como Paramecium, emplean cilios (figura 4-20a y b). Mientras que los flagelos procarióticos giran los eucarióticos se mueven onduladamente (figura 4-2°C). Para ayudar a la expulsión del material extraño de los pulmones las células ciliadas del aparato respiratorio del hombre barren tales materiales a lo largo de la superficie de las células de los bronquios y tráquea hacia la garganta y la boca (ver figura 15-3).
La pared celular
En general, la pared celular eucariótica es considerablemente más sencilla que la de la célula cariótica. La mayoria de las algas poseen paredes celulares formadas por el polisacárido celulosa (como las plantas). Las paredes celulares de algunos hongos contienen también celulosa, pero en la mayoría de ellos el principal componente estructural de la pared celular es el polisacárido quitina, un polímero de unidades de N-acetil-glucosamina (la quitina es también el principal componente de la estructura del exoesqueleto de crustáceos e insectos). La pared celular de las levaduras contiene además los polisacáridos glucano y manano. En los eucariotas que carecen de pared celular la membrana citoplasmática puede ser la cubierta más externa; sin embargo, células como las de los mohos mucilaginosos y amebas, que están en contacto directo con el medio ambiente, pueden presentar otras cubiertas por encima de ella (ver lámina A-30). Los protozoos no tienen una pared celular típica; en su lugar poseen una cubierta externa flexible llamada cutícula.

Figura 4-20. Flagelos eucarióticos y cilios. (a) La euglenoide de agua
dulce Trachelomonas sp. (5.000x). Su largo flagelo en forma de látigo
emerge del cuerpo de la célula en la parte superior de esta fotografía
al microscopio electrónico de barrido. (b) Un ciliado devora a otro
ciliado. Didinium nasatum, la célula inferior con dos franjas de cilios
a su alrededor, ha comenzado a engullir a la célula cubierta de cilios de
Paramecium multimicronucleatum (1.500x). (c) Movimiento de un flajelo
eucariótico.
Una consideración de importancia clínica es que las células eucarióticas no contienen péptido glucano, la base estructural de la pared celular procariota. Su importancia se debe a que algunos antibióticos, como las penicilinas y cefalosporinas, actúan frente al péptido glucano y no afectan, por tanto, a las células eucarióticas humanas.
La membrana citoplasmática
En las células eucarióticas que carecen de pared celular la membrana cítoplasmática es la cubierta externa de la célula. En cuanto a su función y estructura, las membranas citoplasmáticas eucarióticas y procarióticas son muy similares. Sin embargo, existen diferencias en las proteínas. En los eucariotas los azúcares sirven de receptores que realizan funciones como las de reconocimiento célula-célula y permiten también la adhesión de bacterias. Además las membranas citoplasmáticas eucarióticas contienen esteroles, lípidos complejos que no se encuentran en las procarióticas (con la excepción de las membranas de los micoplasmas). Los esteroles parecen estar asociados a la capacidad de las membranas para resistir la lisis debido al incremento de presión osmótica.
Las sustancias pueden atravesar las membranas citoplasmáticas, tanto eucarióticas como procarióticas, por difusión simple, ósmosis, difusión facilitada o transporte activo. En las células eucarióticas no se da la translocación de grupos; en cambio pueden utilizar un mecanismo adicional llamado endocitosis. Para ello una porción de la membrana citoplasmática rodea una partícula o una molécula grande, la encierra y la introduce en la célula. La endocitosis es una de las formas por las que los virus pueden entrar en las células animales.
Dos tipos muy importantes de endocitosis son la fagocitosis y la picnocitosis. En la fagocitosis proyecciones de la célula llamadas seudópodos engloban partículas y las introducen en la célula. Mediante la fagocitosis las células blancas destruyen bacterias y otros cuerpos extraños (véase la figura 15-8 y el capítulo 15). En la picnocitosis la membrana citoplasmática forma invaginaciones que introducen el fluido extracelular en la célula con cualquier sustancia que esté disuelta en él.
Citoplasma
El citoplasma de las células eucarióticas comprende la matriz que hay dentro de la membrana celular y fuera del núcleo (ver figura 4-19). El citoplasma es la sustancia en la que se encuentran diversos componentes celulares. Una diferencia importante con el citoplasma procariótico es que el eucariótico presenta una estructura interna compleja, compuesta de varillas extremadamente pequeñas llamadas microfilamentos y filamentos intermediarios y de cilindros llamados microtúbulos. Conjuntamente forman el citoesqueleto. El citoesqueleto y otros componentes citoplasmáticos se mantienen unidos por una membrana tridimensional de finos filamentos llamada red microtrabecular. Proporciona soporte y morfología, organiza las reacciones químicas que transcurren en el citoplasma y ayuda a transportar sustancias a través de la célula e incluso al movimiento de la célula entera, como ocurre en la fagocitosis. El movimiento del citoplasma eucariótico de una parte a otra de la célula, que contribuye a distribuir nutrientes y al movimiento de la célula sobre una superficie, se denomina corriente citoplasmática. Otra diferencia entre el citoplasma eucariótico y procariótico es que muchas de las enzimas importantes que se encuentran en el fluido citoplasmático de los procariotas se hallan recluidas en los orgánulos de los eucariotas.
Orgánulos
Núcleo
El orgánulo eucariótico más característico es el núcleo (ver figura 4-19). El núcleo suele ser esférico u ovalado (figura 4-21), es frecuentemente la mayor estructura de la célula y contiene casi toda la información hereditaria de la célula (DNA). En las mitocondrias y cloroplastos de los organismos fotosintéticos se encuentra algo de DNA.
El núcleo está separado de la membrana por una doble membrana llamada membrana nuclear. Cada una de estas dos membranas presenta una estructura parecida a la membrana citoplasmática. Diminutos poros de la membrana nuclear permiten la comunicación del núcleo con la red de membranas del citoplasma, llamada retículo endoplásmico (véase el siguiente apartado). Las sustancias que entran y salen del núcleo se cree que pasan a través de estos pequeños poros. Dentro de la cubierta nuclear se encuentra un fluido gelatinoso llamado nucleoplasma; también se encuentran cuerpos esféricos denominados nucléolos. Estas estructuras constituyen un centro para la síntesis de RNA ribosómico, un constituyente esencial de los ribosomas (véase el siguiente apartado).
Finalmente se encuentra el DNA, que aparece combinado con un cierto número de proteínas, entre las que se incluyen varias proteínas básicas llamadas histonas y no-histonas. La asociación de unos 165 pares de bases de DNA y 9 moléculas de histonas constituye un nucleosoma. Cuando la célula no se está reproduciendo, el DNA y sus proteínas asociadas forman una masa con aspecto de ovillo llamada cromatina. Antes de la división nuclear la cromatina se arrolla formando cuerpos más gruesos y en forma de bastoncillos llamados cromosomas. Los nucleosomas son las unidades estructurales básicas de los cromosomas. Los ´cromosomasª procarióticos no llevan a cabo este proceso, no poseen histonas, no están formados por nucleosomas y no están rodeados por una cubierta.
Las células eucarióticas se dividen mediante dos mecanismos complejos llamados mitosis y meiosis (capítulo 8). Ninguno de ellos tiene lugar en las células procariotas.
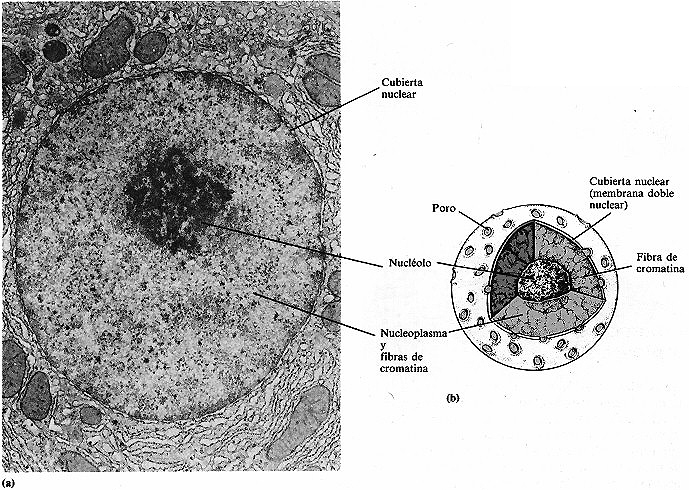
Figura 4-21. El núcleo eucariótico. (a) Fotografía al microscopio
electrónico de transmisión de un núcleo (4.750x). (b)
Esquema de un núcleo.
Retículo endoplásmico
Dentro del citoplasma hay un sistema formado por pares de membranas paralelas que encierran estrechas cavidades de formas variables. Este sistema, que no está presente en procariotas, se conoce como retículo endoplásmico o RE (figura 4-22).
El RE es una red de canales que circulan a través del citoplasma y son continuación tanto de la membrana plasmática como de la membrana nuclear (ver figura 4-19). Se cree que el RE proporciona una superficie para reacciones químicas, una ruta para el transporte de moléculas dentro de la célula y una zona de almacenamiento para moléculas sintetizadas. El RE juega un papel tanto en la síntesis de lípidos como en la de proteínas.
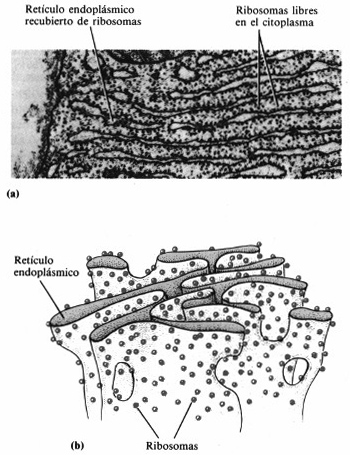
Figura 4-22. Retículo endoplásmico
y ribosomas. (a) Fotografía al microscopio electrónico de transmisión
del reticulo endoplásmico y los ribosomas en sección transversal (24.000x).
(b) Representación tridimensional del retículo endoplásmico
y los ribosomas.