Comprender:
8.1 LAS POBLACIONES
Al estudiar el metabolismo y la genética de los microorganismos, nos hemos centrado en un individuo - la forma en que se mantiene y se reproduce una célula microbiana. En este capítulo y en el siguiente, prestaremos atención a los grandes grupos - estudiando cómo las poblaciones de microorganismos aumentan mediante el crecimiento y disminuyen o desaparecen como resultado de la muerte.
Estudiar las poblaciones requiere que observemos a los microorganismos desde una nueva perspectiva. Cuantío estudiamos individuos, lo que hacemos es describir. Describimos las partes de la célula implicadas en un proceso particular, cómo produce la célula las moléculas que necesita para originar una nueva célula y cómo protege su información genética cuando ésta pasa de una generación a la siguiente. Sin embargo, cuando estudiamos poblaciones, lo que hacemos es Contar. De las poblaciones queremos saber "¿cuánto?","¿cuánto más?"o"¿cuánto menos?". El estudio de las poblaciones, ya sean microbianas o humanas, se realiza desde un punto de vista matemático.
A menos que digamos lo contrario, usaremos el término crecimiento microbiano para referirnos al crecimiento de una población, no al incremento en tamaño de una célula individual. Las células microbianas individuales crecen, pero la mayoria solo incrementan su tamaño hasta alrededor del doble, antes de que se dividan en dos. La división celular se traduce en crecimiento o incremento en el número de celulas de la población (Figura 8.1).
8.2 CÓMO CRECEN LOS MICROORGANISMOS
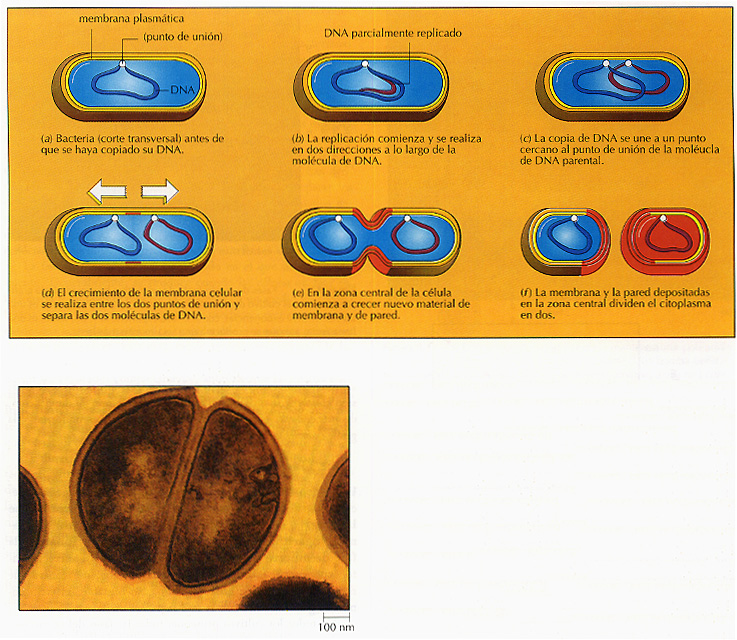
En un ambiente favorable, una célula microbiana dada aumenta su tamaño; y cuando éste se duplica, la célula se divide. La mayoría de las bacterias se alargan y se dividen mediante fisión binaria, o bipartición, cerca del ecuador celular, para formar dos células hijas aproximadamente iguales. Algunos microorganismos unicelulares, incluidas algunas bacterias, se replican por gemación, formando una yema que crece y se separa de la célula parental. Aunque tratamos el crecimiento en términos de una población bacteriana que se divide mediante fisión binaria, los mismos principios se pueden aplicar para los microorganismos que se reproducen mediante gemación (Capítulo 5). El crecimiento de los microorganismos filamentosos (aquellos que forman largos tubos) y de los microorganismos con ciclos de vida complejos siguen reglas más complicadas.
8.2.1 Tiempo de duplicación y tasa de crecimiento
El tiempo de duplicación (también conocido como el tiempo de generación) es el período que requieren las células de una población microbiana para crecer, dividirse, y dar lugar a dos nuevas células por cada una de las que existían anteriormente. El tiempo de duplicación es, aproximadamente, el mismo para todas las células de una determinada población. No cambia hasta que se agotan los nutrientes o comienzan a acumularse los productos metabólicos tóxicos. Cuando las condiciones para el crecimiento se hacen menos favorables, el tiempo de duplicación aumenta antes de que el crecimiento se detenga.
El tiempo de duplicación varía dependiendo de la especie microbiana y de las condiciones de crecimiento de la población. Por ejemplo, el tiempo de duplicación de Escherichia coli es de alrededor de 18 minutos en un medio de laboratorio rico. Pero en el tracto intestinal de los vertebrados, en el cual los nutrientes son menos abundantes, el tiempo de duplicación es de unas 12 horas. Algunas bacterias pueden crecer un poco más rápidamente que E. coli; pero incluso en las condiciones más favorables, muchos microorganismos crecen mucho más lentamente, requiriendo horas o días para duplicar su numero.
El valor del tiempo de duplicación nos indica la rapidez con que está creciendo una población, pero la relación es inversa, Esto es, las poblaciones con un valor de tiempo de duplicación bajo están creciendo rápidamente, mientras que las poblaciones con un valor más elevado de tiempo de duplicación están creciendo más lentamente. Por esta razón, la misma informacion se expresa frecuentemente como tasa de crecimiento, o tiempo de duplicación por hora, que es elevada para las poblaciones de crecimiento rápido y baja para las de crecimiento lento. Así, la tasa de crecimiento de un cultivo de E. coli que tiene un tiempo de duplicación de 18 minutos será de 3,3 (60/18) generaciones por hora.
8.2.2 El crecimiento exponencial
Durante un periodo de tiempo de duplicación constante, una población microbiana se encuentra en crecimiento exponencial. Esto significa que durante cada tiempo de duplicación, el número de células de la población se incrementa en un factor de dos, es decir, se duplica.
El concepto de crecimiento exponencial se ilustra mediante un paradigma matemático acerca de los lirios acuáticos. Un lirio de agua produce una hoja nueva cada día, por cada hoja que tenía
el día anterior. Comenzando con una unica hoja, el lirio cubre un estanque de media hectárea en unos 30 días. Cuánto tardará en cubrir un estanque de una hectárea? La respuesta correcta es 31 días, no 60, porque el tiempo de duplicación es de un día. En condiciones de crecimiento exponencial, los seres vivos se multiplican con una asombrosa rapidez. Durante cada tiempo de duplicación se producen tantas nuevas células como se habían producido anteriormente de manera acumulada.
Figura 8.1 La mayoría de las células microbianas se dividen por fisión binaria como se muestra en esta micrografía electrónica de barrido de la bacteria Staphylococcus aureus. En la Íisión binaria, la célula replica su DNA y proporciona una copia a cada célula hija.
Puesto que el crecimiento microbiano es exponencial, para la expresión gráfica del crecimiento de una población microbiana se utiliza una escala exponencial (Figura 8.2). Aunque la misma información podría expresarse utilizando una gráfica aritmética, o numerica sencilla, la gráfica exponencial tiene ventajas significativas. El numero de microorganismos de una población que aumenta exponencialmente se incrementa lentamente al principio y luego muy rápidamente. Por ejemplo, durante las tres primeras duplicaciones de una sola célula, sólo se producen siete nuevas celulas. Durante la séptima duplicación, se originan 64 células y durante la duplicación vigésimo primera, más de un millón. Si se expresa esta curva en una escala aritmética, el incremento inicial apenas se apreciará, mientras que las fases de crecimiento posteriores dispararían la gráfica. Utilizando una escala exponencial, la curva se convierte en una línea recta, con una pendiente directamente relacionada con la tasa de crecimiento de la población. Esto es lógico si recordamos que por cada duplicación la población se duplica. En otras palabras, el número total de células es igual a dos elevado a un exponente, y dicho exponente es el número de generaciones que han transcurrido desde que la población comenzo a aumentar. Así, una población que comienza como una célula (20), se incrementa a dos células (21), luego a cuatro células (22), ocho células (23), 16 células (24), y asi sucesivamente. La fórmula general es N = 2n, donde N es el número de células en el cultivo después de que pasen n tiempos de duplicación.
¿Por qué la gráfica es una línea recta y no una línea en forma de escalera? Si una población microbiana proviene de una umea célula y tiene un tiempo de duplicación constante, ¿no se mantendría la población constante durante cada tiempo de duplicación y luego se incrementaría bruscamente - en forma de escalera? De hecho, este modelo, denominado crecimiento sincrónico, no se produce en circunstancias ordinarias. Por el contrario, las células individuales de la población suelen dividirse antes o después del tiempo de duplicación de la población, según un modelo de crecimiento asincrónico. Los tiempos de división efectivos se distribuyen alrededor del tiempo de duplicación, de manera que el incremento de la población es uniforme y la gráfica aumenta como una línea recta.
|
aa |
Tiempo | Células / Mililitro | ||
|---|---|---|---|---|
| Notación | ||||
| Minutos | Horas | Número | Científica | Logaritmo |
|
0 |
0 |
1000 |
103 |
3,0 |
|
20 |
0,33 |
2000 |
2 X 103 |
3,301 |
|
40 |
0,66 |
4000 |
4 x 103 |
3,602 |
|
60 |
1,00 |
8000 |
8x103 |
3,903 |
|
80 |
1,33 |
16000 |
1,6X104 |
4,204 |
|
100 |
1,66 |
32000 |
3,2x1O4 |
4,505 |
|
120 |
2,00 |
64000 |
6,4 x 104 |
4,806 |
|
140 |
2,33 |
128000 |
1,28 x 105 |
5,107 |
|
160 |
2,66 |
256000 |
2,56 x 105 |
5,408 |
|
180 |
3,00 |
512000 |
5,12 x 105 |
5,709 |
|
200 |
3,33 |
1024000 |
1,02 x 106 |
6,010 |
(a)
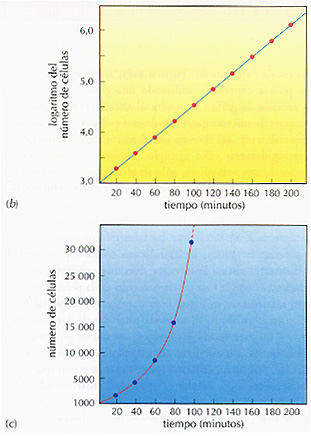
Figura 8.2 Crecimiento exponencial de un cultivo bacteriano. (a) El número de células por mililitro en un cultivo que crece exponencial mente se mide cada 20 minutos y se expresa como el número, de células o como el logaritmo del número de células. (b) La expresión del logaritmo del número de células frente al tiempo es una línea recta. (c) La representación del número de células frente al tiempo es una curva que tiene una pendiente muy acusada. Puesto que el número de células se duplica cada 20 minutos, el tiempo de duplicación es de 20 minutos y la tasa de crecimiento es 3,0 (60/20) duplicaciones por hora. Un cultivo que crece con esta tasa se incrementa, en 200 minutos, desde 1000 hasta más de 1 millón de células por mililitro.
8.2.3 Fases del crecimiento
Un cultivo microbiano pasa típicamente por cuatro fases de crecimiento, distintas y secuenciales: la fase de latencia o fase lag (lag, en inglés, significa "retraso"), la fase log (también llamada logarítmica o exponencial), la fase estacionaria y la fase de muerte (Figura 8.3). Cuando estudiamos el crecimiento exponencial, estamos estudiando la fase log, de manera que retomaremos el tema en este punto. No obstante, el ciclo comienza generalmente con la fase lag.
No todos los cultivos presentan todas las fases del crecimiento. Las fases lag y de muerte pueden existir o no. La fase log tiene lugar casi siempre cuando las células están en un ambiente favorable, pero no puede continuar indefinidamente. De acuerdo con el ejemplo del lirio acuático, la masa de un cultivo microbiano que continuara creciendo de forma exponencial superaría rápidamente el peso del planeta. Pero siempre sucede algo; se agota un nutriente esencial, se acumula un producto tóxico, o el pH se hace desfavorable. Por lo tanto, el cultivo deja de crecer y entra en la fase estacionaria de crecimiento.
Aunque no se produce un aumento neto de la masa del cultivo durante la fase estacionaria, la composición celular cambia cuando el cultivo pasa a esta fase. Las células se hacen más pequeñas y comienzan a sintetizar componentes que les ayudan a sobrevivir, sin crecer, durante largos períodos de tiempo. Por ejemplo, cuando Escherichia coli entra en la fase estacionaria sintetiza alrededor de 30 proteínas, que no se encuentran en las células en la fase log, y modifica la composición de algunos de los ácidos grasos de sus membranas. Las cepas mutantes que no pueden producir estas proteínas de la fase estacionaria mueren rápidamente cuando dejan de crecer. Algunas especies bacterianas han desarrollado sofisticados mecanismos, como la formación de endosporas para sobrevivir a la fase estacionaria (Capítulo 4)
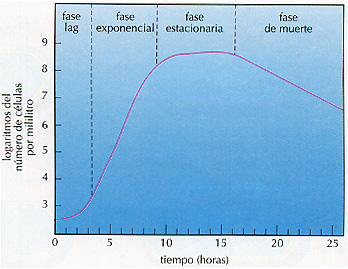
Figura 8.3 Fases de crecimiento de un cultivo bacteriano (esta gráfica se basa en el cultivo descrito en la Figura 8.2). La fase de muerte es una fase de declive exponencial, como la fase exponencial y, por tanto, es una línea recta, aunque se produce a una tasa mucho más lenta.
Después de aproximadamente un día en fase estacionaria, comienza la fase de muerte, y las células del cultivo comienzan a morir. Durante esta fase, las células de la mayoría de las especies mueren exponencialmente, pero a una velocidad lenta - mucho menor que la tasa de incremento de las células durante la fase log. Generalmente, la muerte se produce porque las células han agotado sus reservas intracelulares de ATP y no pueden reparar los componentes celulares. Las células que no crecen no pueden generar nuevo ATP y, por tanto, se produce la muerte cuando no existe suficiente energía para continuar la reparación celular o para reiniciar el crecimiento, cuando existan nutrientes disponibles.
La fase lag es, asimismo, un período en el que no hay crecimiento. Generalmente, se produce cuando las células en fase estacionaria o en fase de muerte se inoculan en un medio de cultivo fresco. Aunque en la fase lag no se produce crecimiento neto, existe una actividad metabólica considerable, ya que las células se preparan para crecer. Esta preparación es necesaria porque los daños metabólicos sufridos durante las fases estacionaria o de muerte deben ser reparados completamente, antes de que las células puedan comenzar a crecer de nuevo. Si las células empleadas para inocular (sembrar) un nuevo cultivo están en la fase log, no existe fase lag, siempre que el nuevo medio de cultivo sea el mismo que el viejo y las demás condiciones permanezcan idénticas.
La duración de la fase lag depende de cuánto tiempo haya estado el cultivo previo en fase estacionaria o de muerte, o puede depender de un cambio en el medio. Si las células han estado en la fase estacionaría sólo brevemente, la posterior fase lag podrá durar sólo unos minutos. Pero si han permanecido en la fase estacionaría durante meses, la subsiguiente fase lag de las células sobrevivientes puede durar muchas horas. Si un cultivo exponencial se pasa de un medio pobre a uno rico, no existe fase lag. Pero el paso de un medio rico a uno pobre implica una fase lag de varias horas, en la mayoría de los cultivos bacterianos.
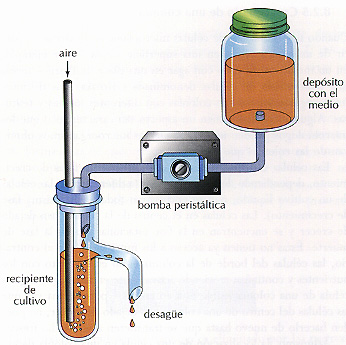
Figura 8.4 El quimiostato se utiliza para mantener una población microbiana en un estado de crecimiento constante. Una bomba peristáltica añade al cultivo el medio estéril desde un reservorio a una tasa constante. El cultivo -que contiene las células y consume el medio parcialmente- sale con la misma tasa. Un quimiostato real tiene muchos controles electrónicos.
8.2.4 Cultivo continuo de los microorganismos
En el laboratorio, la mayoría de los microorganismos se cultivan en sistemas cerrados, lo que significa que no se suministran nutrientes ni se eliminan los productos tóxicos. Consecuentemente, los cultivos pasan, generalmente, por las cuatro etapas de crecimiento. Cuando se inocula un cultivo en fase estacionaria en un medio fresco que contiene las concentraciones adecuadas de todos los nutrientes esenciales, éste pasa secuencialmente por una fase lag, una fase log, una fase estacionaria y una fase de muerte. Sin embargo, los microorganismos no crecen de esta manera en la naturaleza. En la mayoría de las condiciones naturales, los nutrientes entran de manera continua y a bajas concentraciones en el ambiente de una célula, y las poblaciones crecen continuamente a una tasa baja, pero uniforme. La tasa de crecimiento está determinada por la concentración del nutriente limitante, o nutriente más escaso. En la naturaleza raramente la tasa de crecimiento viene fijada por la acumulación de los productos metabólicos, porque generalmente otros microorganismos pueden usarlos a medida que se van formando.
Es posible conseguir un cultivo continuo en el laboratorio en condiciones que imitan las de la naturaleza - manteniendo la concentración de un nutriente esencial lo suficientemente baja como para que limite la tasa de crecimiento. Dicho crecimiento limitado por la concentración se consigue generalmente con un aparato de cultivo continuo denominado quimiostato (Figura 8.4). Un quimiostato consta de una bomba peristáltica, un reservorio, un recipiente de cultivo, y un sistema de desagüe. La bomba añade una cantidad controlada de medio fresco desde el reservorio, de manera continua, al cultivo. Al mismo tiempo, una cantidad idéntica de cultivo sale continuamente a través del sistema de desague. En estas condiciones, el número de células en el vaso de cultivo no cambia. El cultivo crece lo suficientemente rápido como para reemplazar las células que se pierden a través del sistema de desagüe.
La concentración del nutriente limitante en el cultivo fija la tasa de crecimiento del mismo. El cultivo crece a la misma velocidad con que se suministra el nutriente limitante, que es, asimismo, la misma con que se consume parcialmente el medio de cultivo y salen las células mediante el sistema de desagüe. En el vaso de crecimiento del quimiostato, las células permanecen indefinidamente en fase exponencial de crecimiento.
El cultivo continuo tiene aplicaciones industriales importantes (Capítulo 29). Es una forma eficaz de producir células microbianas o sus productos (tales como antibióticos y vitaminas), porque constantemente se obtiene un cultivo denso que sale por el desagüe del vaso de crecimiento.
8.2.5 Crecimiento de una colonia
Cuando una población de células microbianas se desarrolla a partir de una única célula en una superficie sólida - por ejemplo, en un medio solidificado con agar en una placa de Petrí - forma una masa sólida de células denominada colonia. Los distintos microorganismos forman colonias con diferentes formas y texturas. Algunas colonias tienen un aspecto tan característico que los microbiólogos pueden identificar ciertos microorganismos observando las colonias que forman.
Las células de una colonia están en diferentes fases de crecimiento, dependiendo de su localización (a diferencia de las células de un cultivo liquido, que se encuentran todas en la misma fase de crecimiento). Las células en el centro de la colonia han dejado de crecer y se encuentran en la fase estacionaria o en la fase de muerte. Estas no tienen ya acceso a los nutrientes. Por el contrario, las células del borde de la colonia están en contacto con los nutrientes y continúan creciendo activamente en fase log. Ninguna célula de una colonia visible está en fase lag, porque una vez que las células del centro de una colonia han dejado de crecer, no pueden hacerlo de nuevo hasta que se transfieren a un medio fresco.
Asimismo, la localización de una célula en una colonia determina el acceso a los nutrientes de otra forma. Por ejemplo en una colonia que crece en contacto con el aire, las células de la superficie son aerobias. Las células incluidas en el centro de la colonia son totalmente anaerobias, porque las células de la superficie de una colonia que crece en aerobiosis captan todo el oxigeno del aire que difunde a través de la colonia.
8.3 ¿QUÉ NECESITAN LOS MICROORGANISMOS PARA CRECER?
Cada especie microbiana requiere para crecer unas condiciones particulares. Estos requerimientos son muy variados, porque los ambientes en los que las diferentes especies se han adaptado varian enormemente.
8.3.1 Nutrición
Todos los seres vivos utilizan compuestos químicos presentes en el medio ambiente para construir las moléculas necesarias para fabricar nuevas células. Estos compuestos químicos se denominan nutrientes. Sabemos que la nutrición afecta al crecimiento de todos los organismos. Un ser humano, un animal o una planta bien alimentados crecen mas rápidamente que otros mal alimentados. Para los microorganismos, particularmente las bacterias, el efecto de la nutrición en la tasa de crecimiento es enorme. Por ejemplo, Eschenchia coli puede llegar a crecer 10 veces más rápidamente en un ambiente rico en nutrientes, tal como el extracto de carne que en un ambiente nutritivo pobre, como por ejemplo uno compuesto por succinato y sales. El medio con succinato y sales cubre todas las necesidades esenciales de E. coli, pero el crecimiento en dicho medio requiere que la célula realice más procesos biosintéticos. El resultado es un crecimiento más lento.
Una tasa de crecimiento más elevada significa que un cultivo alcanza sus máximos número y masa totales de manera más rápida que en un ambiente pobre en nutrientes. Sin embargo, un cultivo puede alcanzar la misma densidad, al cabo de un tiempo suficiente, siempre que esté presente la misma cantidad total de nutrientes. El rendimiento, o máxima masa total, de un cultivo es directamente proporcional a la cantidad de nutrientes,
Los nutrientes que necesitan los microorganismos para crecer incluyen una fuente de carbono, de oxígeno, de nitrógeno, de fósforo, de azufre, de elementos traza y de factores de crecimiento orgánicos. Las células utilizan estos nutrientes para generar fuerza conductora o metabolitos precursores (Capítulo 5). También se requiere hidrógeno para el crecimiento, pero nunca es un factor limitante en cualquier medio que permita el crecimiento.
Carbono El carbono es la base estructural de los compuestos bioquímicos. Los microorganismos autotrofos obtienen su carbono a partir del dióxido de carbono (C02) de la atmósfera, mientras que los microorganismos heterotrofos obtienen el carbono a partir de los compuestos orgánicos del ambiente (Capítulo 5).
Los microorganismos heterotrofos utilizan como fuente de carbono muchas moléculas orgánicas diferentes. De hecho, probablemente cualquier compuesto orgánico que se encuentra en la naturaleza es utilizado como fuente de carbono por alguna especie bacteriana. Una de las fuentes más comúnmente utilizada por los microorganismos es la glucosa, una hexosa que juega un papel central en el metabolismo. Consecuentemente, la glucosa es un ingrediente común de los medios de cultivo microbiológicos. No obstante, frecuentemente se añaden a los medios como fuente de carbono otros azúcares, así como polisacáridos, ácidos orgánicos, alcoholes y aminoácidos.
Para los quimioheterotrofos, las moléculas orgánicas que contienen carbono, como la glucosa, proporcionan tanto una fuente de energía para generar ATP, como los átomos de carbono necesarios para fabricar los compuestos bioquímicos (Capítulo 5). Las fuentes de carbono que son metabolizadas rápidamente permiten un crecimiento relativamente rápido, mientras que las fuentes de carbono que son metabolizadas más lentamente permiten sólo un crecimiento lento. Por ejemplo, en un medio de cultivo al que se añade glucosa como fuente de carbono en lugar del aminoácido lisína, Escherichia coli crece el doble de rápido que en un cultivo idéntico excepto la adición mencionada.
Oxígeno Todos los microorganismos requieren oxígeno elemental para fabricar sus componentes bioquímicos, pero no todos los microorganismos requieren oxígeno atmosférico. La mayoría de los microorganismos heterotrofos obtienen el oxígeno a partir de la misma molécula que les sirve como fuente de carbono. La fórmula química de los hidratos de carbono (una fuente de carbono común para los heterótrofos) es CH2O, que significa que cada molécula de hidratos de carbono proporciona a la célula un átomo de oxígeno por cada átomo de carbono. Los autotrofos, que generan energía a partir de la luz, obtienen la mayor parte de su oxígeno a partir del CO2 fijado durante la fotosíntesis. La mayoría de los microorganismos aerobios tienen enzimas denominadas oxigenasas que pueden añadir directamente oxígeno atmosférico a las moléculas orgánicas, pero ésta es una fuente minoritaria de oxígeno. Algunas enzimas permiten a los microorganismos utilizar el oxigeno presente en el agua mediante reacciones hidrolíticas.
Redescubridores de microbios
Los supervivientes
¿Cómo pueden sobrevivir las bacterias no formadoras de endosporas cuando dejan de crecer? Unas pocas horas después de que Escherichia coli (y otras bacterias no formadoras de endosporas) entra en la fase estacionaria de crecimiento, comienza la fase de muerte. Aunque el declive de la fase exponencial puede ser muy lento, puede ocurrir la muerte de todas las células del cultivo. Según esto, ¿sería sólo una cuestion de tiempo la eliminación de todas las especies bacterianas? De hecho, esto no ocurre porque no todas las células mueren durante la fase de muerte. Incluso después de un año de almacenamiento, un cultivo de E. coli en fase estacionaría contiene más de un millón de células vivas por mililitro. Las células del cultivo mueren exponencialmente en cuatro o cinco días, pero las células que permanecen sobreviven durante largos períodos de tiempo. Roberto Kolter, un microbiólogo de la Universidad de Harvard, encontró que las bacterias no formadoras de endosporas han desarrollado sus propios mecanismos para sobrevivir períodos prolongados de escasez de nutrientes. La mayoría de las células mueren, pero unas pocas están programadas para sobrevivir, lo cual es suficiente para preservar la especie.
Además de utilizar el oxígeno como nutriente, los microorganismos que son capaces de una respiración acróbica usan el oxígeno para generar energía. Pero, para algunos organismos el oxigeno atmosférico es tóxico. Algunas enzimas son destruidas rapída e irreversiblemente por la exposición al oxígeno atmosférico. Un ejemplo extremo es la nítrogenasa, enzima que permite a las bacterias fijadoras de nitrógeno utilizar el nitrógeno atmosférico. Como resultado, los microorganismos han desarrollado varios mecanismos para proteger incluso las enzimas altamente sensibles al oxigeno y, por tanto, sobrevivir en presencia del oxígeno. Algunas cianobacterias fijadoras de nitrógeno - que son acróbicas y producen oxígeno gaseoso mediante fotosíntesis - protegen la nitrogenasa, segregándola en células especializadas, denominadas heterocistes. Los heterocistes no producen oxígeno y son impermeables al mismo. La bacteria fijadora de nitrogeno Azotobacter realiza una respiración aerobica a una tasa lo suficientemente evada como para mantener anaeróbico el centro de la célula, donde está localizada la nítrogenasa.
Además del oxígeno, los organismos deben hacer frente a los compuestos tóxicos que contienen oxigeno y que se producen durante el metabolismo acróbico. Entre estos agentes tóxicos se encuentran el peróxido de hidrógeno (H2O2) y los radicales libres, que son incluso más tóxicos: superóxido (O2) y el radical hidroxilo (·OH), que se forma a partir del superóxido. Un radical libre es un compuesto con un electrón sin aparear que lo hace extraordinariamente reactivo. Si se acumula superóxido, los compuestos bioquímicos se oxidan rápidamente y las células mueren.
Los seres vivos que sobreviven en presencia del aire han desarrollado sistemas para eliminar los radicales libres. Todos los aerobios producen la enzima superóxido dismutasa, que destruye el superóxido convirtiéndolo en oxígeno y peróxido de hidrógeno
![]()
El peróxido de hidrógeno, que es el producto de ésta y de otras reacciones catalizadas por enzimas, es en si mismo un oxidante tóxico. Se convierte en agua y oxígeno mediante la enzima catalasa.
![]()
El peróxido de hidrógeno también es eliminado por las peroxidasas, enzimas que lo utilizan para oxidar ciertos compuestos orgánicos. Los organismos que pueden tolerar la exposición al oxigeno contienen superóxido dismutasa, y casi todos también contienen catalasa. Los organismos que pueden tolerar la exposición al oxígeno contienen superóxido dismutasa, y casi todos también contienen catalsa. Los organismos que carecen de ambos enzimas mueren rápidamente en un ambiente con oxígeno.
La relación entre el oxígeno y el crecimiento varia enormemente de una especie microbiana a otra. Los aerobios obligados pueden crecer tan solo en presencia de oxígeno porque para generar energía requieren oxígeno como aceptor terminal de electrones. Los microaerófilos toleran menos oxígeno (o requieren eoncentraciones mayores de CO2). Éstos pueden crecer sólo a concentraciones reducidas de oxigeno (del 2 al 10 por ciento de oxígeno, en comparación con el 21 por ciento de oxígeno presente en el aire). Los anaerobios facultativos pueden crecer tanto en presencia como en ausencia de oxígeno, porque cuando éste está disponible, lo utilizan como aceptor terminal de electrones, pero cuando el oxigeno está ausente, utilizan otras rutas bioquímicas para generar energía. Los anaerobios aerotolerantes pueden generar energía sin oxígeno pero no mueren al ser expuestos al aire. Los anaerobios obligados mueren en presencia del oxígeno, porque carecen de los enzimas necesarios para eliminar el superóxido (Tabla 8.1).
Nitrógeno El nitrógeno constítuye alrededor del 14
por ciento del peso seco de la mayoría de los microorganismos. Es un constituyente
de las proteínas y de los ácidos nucleicos así como de algunos
metabolitos esenciales, de manera que todos los seres vivos requieren una fuente
de este elemento. La forma de nitrógeno que utilizan los microorganismos depende
de su capacidad metabólica y del ambiente en que se encuentran. Probablemente,
todos los microorganismos pueden usar amoníaco (NH3) como fuente
de nitrógeno, porque esta forma es la que se incorpora en la biosíntesis.
La mayoría de los microorganismos pueden obtener amoníaco a partir
de una variedad de compuestos orgánicos, incluidos los aminoácidos,
y algunos pueden utilizar formas inorgánicas, entre las que se incluye el
ion nitrato (NO3). Algunos pueden lijar el nitrógeno atrnosférico gaseoso
(N2); estos organismos lijadores de nitrógeno prosperan en los ambientes en
los que el nitrógeno es el nutriente limitante.